Анастасия Волчок, Валерия Ню
«Наука из первых рук» №1(77), 2018

Уже давно растениеводство во всем мире радуется многочисленным подаркам генной инженерии — устойчивым к вредителям и холоду, быстрорастущим и продуктивным растениям, однако для среднестатистического потребителя метка «ГМО» сегодня сродни печати Каина. Почему так? Один ответ лежит на поверхности: генные инженеры делают сегодня то, чего природа не смогла бы сделать никогда или, по крайней мере, очень нескоро, что не только впечатляет, но и пугает. Людей настораживает и то, что сами создатели ГМО не торопятся полностью исключать риск возможных негативных последствий их распространения. С другой стороны, согласно свежим опросам «Левада-центр», всего лишь 30% россиян точно знают, что гены содержатся во всех растениях, а не только в генетически модифицированных, поэтому тотальная ГМО-боязнь во многом вызвана тотальной «генетической» безграмотностью. Между тем спектр современных методов селекции новых сортов растений очень широк, а среди разрешенных есть не менее рискованные и/или практически неизвестные широкой общественности подходы.
Об авторах
|
Анастасия Александровна Волчок — кандидат химических наук, младший научный сотрудник лаборатории биотехнологии ферментов ФИЦ «Фундаментальные основы биотехнологии» РАН (Москва). Победитель весеннего финала «У.М.Н.И.К» Московского государственного университета им. М. В. Ломоносова (2013). Автор и соавтор 11 научных публикаций и 1 патента. |
|
Валерия Анатольевна Ню — специалист по качеству FMCG-товаров и безрецептурных лекарственных средств в центральном офисе компании «Джонсон & Джонсон» (Москва). |


ГМО расшифровывается как генетически модифицированный организм, что подразумевает то или иное воздействие на геном растений — хранилище наследственной информации, «святую святых» живых клеток. Сегодня большинство новейших методов селекции растений тяготеют к внесению изменений непосредственно в структуру ДНК. В результате, по определению ВОЗ, мы и получаем ГМ-растения — новые сорта, которые не могли образоваться в природе в результате размножения или естественной рекомбинации («перемешивания генов»).
Все это так, но в изменении генома растений на самом деле нет ничего нового и революционного! Человек всегда так или иначе оказывал влияние на генетику растений, которые выращивал, хотя и понятия не имел о генах.
Путь к современным культурным растениям, которые мы употребляем в пищу, начался примерно 10 тыс. лет назад, когда появилось сельское хозяйство. Человек выбирал самые здоровые и пригодные для еды растения и планомерно их выращивал. В сельском хозяйстве нет места закону естественного отбора: согласно закону человека (искусственному отбору), выживает только то растение, которое отвечает его запросам.
Ярким примером расхождения целей естественного отбора и селекции служит кукуруза. У предка этого злака зерна при созревании легко отделялись от початка и падали на землю. Такая кукуруза прекрасно размножалась, но человек неизбежно терял большую часть урожая. Что же мы видим теперь? Ядра современной кукурузы на момент зрелости прочно прикреплены к початку. Так же обстоит дело и с другими зерновыми культурами — рисом, ячменем, пшеницей.

Все эти новые виды культурных растений, по сути, являются результатом модификации генома разными способами, например, путем скрещивания разных сортов, что приводит к появлению совершенно новых культурных форм. Огромный материал для искусственного (так же как и для естественного) отбора предоставляет природный мутационный процесс. Ведь спонтанные мутации (изменения) в ДНК растений происходят постоянно, например, в результате действия солнечного излучения. И если такая мутация приводит к появлению особей с заметными положительными отличиями, их остается только тиражировать — вот и вся селекция. Примером служит большое разнообразие современных овощей семейства крестоцветных: брокколи, цветная и белокочанная капуста происходят от одного общего предка (Kempin et al., 1995).
Дальше — больше. За последние 80 лет люди получили более 3 тыс. новых сортов растений, воздействуя на исходные формы излучением или химическими реагентами, чтобы вызвать непредсказуемые мутации в ДНК. Растения, полученные в результате такого искусственно вызванного ненаправленного мутагенеза, успешно возделывают и поныне. Более того, как это ни парадоксально, они никогда не считались ГМО. Впоследствии в обществе распространилось крайне ошибочное мнение, что первые генетически модифицированные растения появились лишь в результате использования методов генной инженерии, целенаправленно воздействующих на ДНК.
В любом случае оценивать новый сорт следует исходя из его характеристик, а не того или иного пути селекции. А чтобы составить собственное мнение об опасности ГМО, нужно как минимум понимать, откуда они берутся.
Рецепт ГМО: режь, исправляй, сшивай
Так как же сделать ГМО из обычного растения? Рецепт довольно прост. Возьмите геном растительной клетки и добавьте в него «генетическую конструкцию» — последовательность ДНК, кодирующую производство нужного белка. Доставить ген можно с помощью вектора — молекулы ДНК или РНК, способной «размножаться» и переносить чужеродный наследственный материал из клетки в клетку. Таким образом можно добавить сразу несколько генов, например, с помощью вектора на основе кольцевой бактериальной плазмиды.

На первый взгляд, все просто, если не задумываться о том, как вставить новый генный фрагмент именно в тот участок ДНК растительной клетки, который нам нужен. А ведь в этом и заключается самая сложная задача редактирования генома, результатом которого являются современные ГМО.
Для того, чтобы расщеплять молекулы ДНК в точно заданных участках, сначала стали использовать рестриктазы — ферменты-«ножницы», способные узнавать определенные последовательности нуклеотидов (строительных блоков ДНК). Функцию сшивки ДНК-цепи исполняли другие ферменты — ДНК-лигазы, призванные исправлять (репарировать) повреждения в структуре ДНК.
Сегодня, как и 30–40 лет назад, эти методы активно используют для получения новых вариантов бактериальных и вирусных геномов. А вот для успешной работы с геномами высших организмов (таких как растения, животные и мы с вами) этих инструментов оказалось недостаточно. Дело в том, что рестриктазы способны узнавать лишь короткие последовательности ДНК, что вполне достаточно для эффективного расщепления коротких ДНК-цепей бактерий, где такие участки встречаются нечасто. Но геномы высших организмов содержат огромное множество коротких последовательностей нуклеотидов, узнаваемых рестриктазами, поэтому «прицельность» метода оказывается очень плохой.
Для редактирования таких геномов пришлось создавать свои инструменты точечного воздействия на ДНК: сначала олигонуклеотид-направленный мутагенез растений, затем сайт-направленный мутагенез с использованием ферментов-нуклеаз с «цинковыми пальцами», TALENs-нуклеаз и даже мегануклеаз (Закиян, 2014; Daboussi, 2015). Но лишь с открытием в 2012–2013 гг. знаменитой технологии CRISPR/Cas9 ученые вплотную подошли к точному исправлению или редактированию генов и геномов (Cong, 2013). Возможность вносить контролируемые изменения в наследственную информацию живых клеток стала настоящим прорывом и повлекла за собой глобальные изменения в селекции.
Насколько остры генетические ножницы?
Основой системы CRISPR/Cas9 стал своеобразный молекулярный механизм, с помощью которого бактерии защищаются от бактериофагов (бактериальных вирусов). При проникновении патогенного вируса в бактерию запускается «иммунная» реакция, приводящая к расщеплению чужеродной генной последовательности. Это делает белок-«ножницы» Cas после того, как захватчик распознан по генному «портрету» — фрагментам вирусной ДНК, хранящимся в участке CRISPR бактериального генома.
На основе бактериальных CRISPR/Cas-систем ученые создали упрощенные искусственные молекулярные конструкции, включающие белок Cas9 и обеспечивающие невероятную точность при разрезании цепей ДНК (Закиян, 2014). С их помощью стало возможным проводить все виды модификаций генома: вносить точечные мутации, встраивать, исправлять, заменять или удалять крупные ДНК-последовательности и фрагменты выбранных генов.

С помощью системы CRISPR/Cas9 уже внесены точные модификации в геном множества растений, в частности, получены новые сорта риса, устойчивые к поражению гнилью, вызываемой фитопатогенными бактериями Xanthomonas, а также знаменитый «золотой рис», содержащий ген бета-каротина (Chen, Gao, 2013). Была решена и нетривиальная задача — создание растений-«биофабрик», способных синтезировать белки человека: инсулин, необходимый для больных сахарным диабетом, и альбумин, применяемый при лечении ожогов и цирроза.
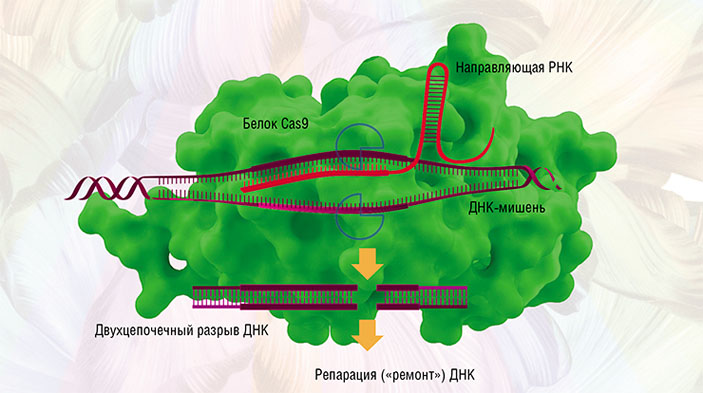
Но несмотря на подтвержденную эффективность системы CRISPR/Cas9 все еще остается риск неспецифичного воздействия на ДНК и нарушения последовательности кодирующих генов. Неудивительно, что настоящий взрыв в мировом сообществе вызвала публикация китайских ученых из Университета Сунь Ятсена (КНР), несколько лет назад впервые применивших CRISPR/Cas9 для исправления генома эмбрионов человека с целью лечения генетического заболевания талассемии. При этом лишь для 4-х из 86 подопытных оплодотворенных яйцеклеток удалось достичь положительного результата (Liang et al., 2015).

Сегодня ряд специалистов призывают к мораторию на любые эксперименты, связанные с редактированием генов человеческих эмбрионов или половых клеток. Их опасения можно понять: когда речь идет о геноме человека, успех должен быть гарантирован. И все же прогресс не остановить: недавно Великобритания стала второй страной, где исследователям было позволено проводить подобные эксперименты (Ершов, 2016).
Тем не менее страх человека перед вмешательством в геном живых организмов не только не убывает, но и в некоторых случаях даже продолжает расти. Вследствие этого оборот и потребление продуктов геномного редактирования растений строго контролируются на законодательном уровне, что препятствует переходу мирового сельского хозяйства на использование продвинутых методов селекции. Однако ученые не сдаются и предлагают сократить до минимума и даже исключить возможные риски негативных последствий введения новых генов в организм растений.

Снижаем риски: от ТРАНС к ЦИС и ниже
Сейчас прохождение всех тестов на биобезопасность и вывод на рынок генетически модифицированных организмов, в том числе растений, жестко регулируется на международном уровне. В этом вопросе правовая база ЕС опирается на директиву Европейского парламента и Совета Европейского союза от 12 марта 2001 г. «О преднамеренном выпуске в окружающую среду генетически модифицированных организмов». Примечательно, что данный нормативный документ исключает из перечня ГМО организмы, полученные путем скрещивания, экстракорпорального оплодотворения, полиплоидной индукции, возникновения мутаций и слияния протопластов скрещиваемых видов (соматическая гибридизация).
Законодательство РФ в области ГМО растительного происхождения регулируют 4 федеральных закона и 6 постановлений Правительства РФ, в том числе федеральный закон № 86-ФЗ «О государственном регулировании в области генно-инженерной деятельности» от 5 июля 1996 г. Ожидается вступление в силу постановления, позволяющего узаконить возделывание ГМ-культур на территории России, которые пока можно выращивать только на опытных участках. Для ввоза в нашу страну сегодня разрешены 22 линии пищевых и кормовых ГМ-растений, среди которых кукуруза, картофель, соя, сахарная свекла и рис, а любые ГМО и ГМ-продукты должны проходить обязательную регистрацию.

В свою очередь, мировое ученое сообщество считает, что нужно различать ГМО по способу получения и делать послабления для продуктов, полученных умеренным вмешательством. Так появилась система деления ГМО на три вида: ТРАНС, ЦИС и ИНТРА.

Трансгенными сегодня называют организмы с искусственно введенными генами, которые в принципе не могут быть приобретены путем естественного скрещивания. Это могут быть гены растений других видов или животных, например рис, в геном которого встроен ген кукурузы. Потенциальная опасность трансгенных культур в том, что приобретенные таким образом новые качества могут повлиять на пригодность к использованию в пищевых или кормовых целях, а затем передаться диким родственникам, что может иметь непредсказуемые последствия для природных экосистем. По этой причине законодательные и регулирующие органы развитых стран уделяют большое внимание биобезопасности таких культур, чтобы снизить риск экологических сдвигов.

![]()
В геном цисгенных растений могут быть введены гены организмов того же или близких видов, с которыми возможно скрещивание в естественных условиях. При этом сам целевой ген не должен быть видоизменен или оторван от своих регуляторных последовательностей. Пример цисгенного растения — картофель, не подверженный картофельной гнили благодаря встраиванию генов диких видов картофеля из Анд, устойчивых к этому заболеванию. Такой картофель сейчас создается в Бельгии (VIB’s fact series, 2015). Важно, что цисгенезис не привносит в организм растения принципиально новых для него признаков и, по сути, аналогичен традиционному скрещиванию с родственными дикими формами.
Интрагенезис можно считать продолжением концепции цисгенезиса, но в этом случае в ДНК растения встраивают его собственный ген, совмещенный с регуляторными участками других его генов. В ходе такой модификации искусственно создаются новые комбинации из уже имеющихся в растении участков ДНК (Holme, 2013). Подобное изменение регуляции активности генов позволяет усиливать полезные признаки (например, способность накапливать витамины в листьях) или, напротив, устранять или сводить к минимуму нежелательные.
Между тем при современном регулировании оборота ГМО-различия между трансгенными и цисгенными растениями не учитываются, хотя эти типы кардинально различаются. Из-за жестких рамок, установленных законодательством, получение и использование цисгенных растений серьезно затруднено, что может заблокировать или значительно отсрочить проведение дальнейших исследований по улучшению сортов сельскохозяйственных культур. Пока лишь в Канаде контроль за цисгенными растениями менее строг по сравнению с трансгенными (Schouten, 2006).
Соматический Франкенштейн
Интересно то, что в мощной законодательной «обороне» против ГМО оказались бреши, появившиеся благодаря ряду парадоксов и допущений, которые на руку смелым селекционерам. Один из примеров — соматическая гибридизация. Другими словами, формирование новых форм растений путем комбинирования ядерных и других (митохондриальных и пластидных) генов при культивировании и слиянии обычных соматических клеток, составляющих ткань растения и не принимающих участия в половом размножении. Этот тип гибридизации растений достаточно широко распространен, при этом на территории ЕС такие соматические гибриды не считаются ГМО. Соответственно их оборот не подвергается строгому контролю.
Что же это за волшебный способ селекции? На первом этапе клетки растений двух разных видов (как правило, культурного и дикорастущего) обрабатывают специальными агентами, разрушающими клеточную оболочку, чтобы получить протопласты. Далее химическим или механическим способом провоцируют слипание и слияние протопластов, которые в дальнейшем восстанавливают общую клеточную оболочку. В результате из двух и более «родительских» клеток образуется новый живой организм — регенерант, или соматический гибрид.
Судьба родительских геномов при этом может быть различной. Два ядра могут синхронно делиться без слияния, образуя двуядерные дочерние клетки. Если же они сольются во время митотического деления, то в итоге получатся устойчивые одноядерные дочерние клетки, несущие смешанный генетический материал. Что касается внеядерного генома, то он тоже может быть получен как от одного родителя, так и быть смешанным. С помощью соматической гибридизации можно получать самые разные гибриды, включая такие, создание которых в принципе невозможно половым путем: например, гибриды, несущие цитоплазматические гены не от материнского растения, а от обоих родителей; «цибриды», содержащие ядро от одного из родителей, а цитоплазму от другого, и др.

Использование соматических клеток при гибридизации позволяет успешно работать с отдаленными, обычно нескрещиваемыми видами и полностью стерильными растениями. Иными словами, этот метод используют, если возникает необходимость преодолеть несовместимость культурных и дикорастущих видов. Таким способом можно получать межклассовые гибридные клеточные колонии: рис + соя, ячмень + табак и даже табак + мышь (Makonkawkeyoon, 1995)! Правда, большинство таких регенерантов сами размножаться уже не способны, а иногда и вовсе представляют собой скорее скопление клеток, чем полноценный организм.
Интересно, что, хотя метод соматической гибридизации влечет за собой значительную «перетасовку» генов, а его результаты очень непредсказуемы, он, тем не менее, разрешен для использования в сельском хозяйстве, в отличие от методов направленного мутагенеза. Как говорила Алиса в Стране чудес, «чем дальше, тем страньше».
Что скрывается под прививкой
А теперь пришло время обратиться к методам, которые должны удовлетворить стандарты даже самых ярых приверженцев натуральных продуктов. Ведь методы эти используются уже очень давно, и они не встречают общественного или законодательного сопротивления. Но оказывается, что с точки зрения генетики эти методы вовсе не «безгрешны», а их комбинация с новейшими подходами открывает перспективы, о которых вы, возможно, и не подозревали.
Например, давно известным способом размножения растений, с которым повсеместно сталкиваются садоводы-любители, является прививка. Суть ее в том, что стебель одного растения (привой) пересаживают на корень другого (подвой). В конце XIX в. этот метод помог спасти европейские сорта винограда Vitis vinifera от нашествия насекомого филлоксеры, повреждающего корни. Прививку осуществляли на североамериканский виноград Vitis labrusca, устойчивый к этому вредителю (Трошин, 1999). А в 2003 г. фермер из Орегона Р. Баур с помощью прививки получил настоящий «томак» (томат + табак), совсем как у Гомера из популярного американского мультсериала «Симпсоны». Пробы показали наличие в томаке никотина, правда, только в листьях, а не в плодах (Philipkoski, 2003).
Что же происходит с растением в результате прививки, если геномы подвоя и привоя не меняются? Во-первых, у подвоя могут увеличиваться или уменьшаться сила роста, размер плодов, сроки созревания. При этом новые признаки не передаются потомству в случае размножения семенами, так как не являются наследственными. Во-вторых, в результате использования прививки возможны «химерные» изменения, и в итоге привитое растение будет состоять из генетически разнородных клеток. Этот эффект также не наследуется. Его часто используют в декоративном цветоводстве и садоводстве для получения растений с мозаичной окраской листьев или соцветий.

В ходе прививки возможно и появление настоящих мутаций, спровоцированных специфическими веществами (этилметансульфонатом, этилимином и др.), которые поступают к привою от подвоя. Однако частота появления мутаций после прививок крайне низка. Неоспоримым преимуществом прививок является возможность размножать мутации, не передающиеся по наследству, а основным недостатком — большой объем исходного материала.

Прививка растений — это, безусловно, метод проверенный и безопасный. Но что произойдет, если в качестве подвоя использовать растение, перенесшее генетическую модификацию? Будет ли полученное растение ГМО? Оказывается, нет: согласно законам, плоды таких гибридов не входят в перечень ГМО, так как ДНК привоя остается неизмененной. Однако мы не можем быть уверены в том, что никакого обмена наследственной информацией между привоем и подвоем не происходит. К примеру, от корневища к привою могут перейти молекулы РНК, регулирующие работу генома, а это означает, что нельзя предсказать и уровень производства тех или иных белков в привитом растении.
Берем генетический разбег!
Но прививка — это далеко не единственный окольный путь для создания новых сортов с измененной активностью ДНК. Ускоренное скрещивание деревьев и кустарников (fast-track breeding) — это даже не метод, а целый комплекс методик, направленных на сокращение сроков получения новых сортов, что особенно важно для многолетних культур. Ведь цикл размножения деревьев с крупными плодами (например, ореха или сливы) может доходить до 10 лет и более (van Nocker, 2014). Это означает, что после посадки первого гибрида селекционер вынужден ждать 5–10 лет, пока тот вырастет и повзрослеет, чтобы продолжить работу. Если же необходимо провести несколько последовательных скрещиваний, выведение нового сорта дерева может занять и 30 лет. В современных условиях никто не готов столько ждать.

Чтобы максимально ускорить процесс, ученые давно поливают своих подопытных гормонами роста, выращивают их при высоких температурах и прибегают к другим уловкам, таким как ДНК-технологии. Среди безобидных можно отметить маркер-вспомогательный отбор, который заключается в анализе генома новых ростков или даже семян и отборе лучших гибридов задолго до того, как они превратятся во взрослые растения. Теперь растение уже не нужно обрабатывать патогеном, чтобы понять, насколько оно к нему устойчиво, достаточно найти нужный ген в семечке. Основной недостаток такой селекции — ее высокая стоимость, поскольку скрининг ДНК — вещь недешевая.
Чтобы растение быстрее повзрослело, селекционеры иногда хитрят. Например, искусственно активируют гены, отвечающие за запуск механизма размножения, после чего начинает цвести и приносить плоды совсем молодое растение. Иногда в геном дерева вводят дополнительные гены, которые ускоряют процессы цветения и плодоношения, и время ожидания первого цветения саженцев сокращается до 1 года. При сочетании методик ускоренного и возвратного (когда гибрид скрещивают с одним из родителей) скрещиваний ген быстрого цветения можно сначала ввести в исходный сорт, а на последнем этапе селекции удалить его путем скрещивания генетически измененного гибрида с родительским растением.
Ускоренное скрещивание осуществляют также путем прививания на ГМ-подвой. Секрет в этом случае кроется в генетически измененном корневище, в котором активно работают гены, отвечающие за цветение. В результате из корневища к листьям поступают специфические белки, запускающие механизм взросления, и привой начинает цвести.
Таким образом, современные методы прививки и ускоренного скрещивания растений за внешней традиционностью таят в себе много настоящих генетических секретов. В то же время ученые, сталкиваясь с общественным мнением и жестким регулированием распространения ГМО, все чаще пытаются избежать внесения изменений непосредственно в растительную ДНК. И здесь мы вплотную подходим к самой загадочной группе современных методов селекции.
Эпигенетика: чуть-чуть не считается
Одними из самых молодых и суперсовременных альтернатив для селекционеров стали подходы эпигенетики — науки о наследуемых механизмах управления экспрессией генов (Marjori, 2015). Как работает наш генетический код, общеизвестно, но вот тонкости его надстройки (эпи- означает ‘над’), своего рода «дирижера», управляющего работой генома, во многом остаются загадкой.
Начало производства белков в клетке регулируется множеством факторов. У клетки также имеются приемы, заставляющие «замолчать» тот или иной ген, чтобы предотвратить производство уже ненужного белка: это и разрушение еще незрелых молекул РНК, считанных с генетической «матрицы», и создание «механических» препятствий для самого считывания ДНК (Marjori, 2014). В общем, эпигенетических сигналов в клетке очень много, они не до конца изучены, однако некоторые из них уже используются для селекции растений, которые попадают к нам на стол каждый день.
Подавить работу генов в клетке можно с помощью природного механизма — РНК-зависимого ДНК-метилирования, суть которого состоит в присоединении метильной группы (СН3) к нуклеотиду цитозину, стоящему в определенном положении. В результате блокируется процесс считывания информации с ДНК на молекулу РНК (Zhang, 2013).

Метилирование ДНК у растений и животных осуществляется ферментами ДНК-метилтрансферазами. Сами по себе эти ферменты метилировать ДНК не могут: им нужны специальные некодирующие РНК, которые направляют метилтрансферазы к конкретным участкам ДНК. Более того, считается, что в метилировании ДНК участвуют еще два вида РНК: малые интерферирующие РНК и микроРНК. Все вместе эти молекулы и определяют, какой именно участок ДНК цепи нужно метилировать. Сегодня такие РНК можно ввести в растение с помощью разных методик, например, посредством вирусов растений или с помощью техник генной инженерии (Deng, 2014).
Интересно, что если ученый изменяет признак растения с помощью ДНК-метилирования и при этом не вносит в геном никаких мутаций, то такое растение не считается ГМО. Если же некодирующие РНК не вводятся извне, а производятся самим растением благодаря геномному редактированию, то оно уже относится к генно-модифицированному.
Но и тут можно схитрить. Дело в том, что у растений метилирование определенных областей ДНК может наследоваться, т. е. передаваться от родителей к следующим поколениям (Jones, 2001). Благодаря ряду скрещиваний ГМ-растения с его природной формой можно получить гибрид, у которого нет измененной ДНК, но метилирование сохраняется. Такой гибрид уже не будет считаться генетически модифицированным.

Насколько метилирование безопасно? Достаточно, ведь метильные группы присоединяются к ДНК совсем не в случайных местах. Поэтому, в отличие от традиционных методов селекции, результаты такого воздействия предсказуемы: мы можем заранее выбрать ген, кодирующий определенный белок, и просто заставить его замолчать. Но делать это нужно аккуратно, так как механизмы метилирования ДНК довольно сложны. Иначе в результате мы можем получить растение, подверженное болезням или преждевременному старению.
Иногда ДНК-метилирование, наоборот, является обязательным условием для начала работы гена. Ученые и это научились использовать: с помощью изменения метилирования ДНК можно увеличить активность генов, отвечающих за производство растением запасных белков. Например, регулируя метилирование, можно повысить содержание белков в зерне пшеницы, а путем обработки риса ингибитором метилирования (5-азацитидином) — получить растения с наследуемым признаком карликовости (Ванюшин, 2013).
Молчание РНК как заслон от аллергии
Успешное считывание гена на матричную РНК вовсе не означает, что кодируемый им белок будет построен: эта мРНК может быть разрушена в цитоплазме клетки. Такое явление, названное посттранскрипционным молчанием, часто наблюдается при внесении дополнительных генов в ДНК растений. Впервые оно было описано еще в 1990 г., когда при введении в геном петунии дополнительных копий гена, отвечающего за красную окраску цветков, количество красного пигмента не только не возросло, но и значительно снизилось (Napoli et al., 1990).

Механизм «замолкания РНК» снижает эффективность работы генных инженеров. С другой стороны, его можно использовать для создания растений, устойчивых к растительным вирусам, так как он может способствовать разрушению не только их собственных матричных РНК, но и соответствующих РНК вирусов, которым удалось проникнуть в клетки растений.
Судя по всему, механизмов посттранскрипционного молчания существует несколько, и ученые пока не вполне понимают, как они работают и как связаны между собой, белые пятна в этой области только начинают заполняться (Плотников, 2007). Есть предположение, что отдельные молекулы мРНК активно деградируют при достижении определенного порога своей численности (Abler, 1996). Другая теория основана на изменениях в регуляции работы генома, связанных с метилированием ДНК, в результате чего среди нормальных РНК синтезируется некоторое количество «ненормальных», которые и запускают распад мРНК в цитоплазме (Hoofvan, 1997).
Одним из хорошо описанных механизмов посттранскрипционного молчания является РНК-интерференция. Этот метод базируется на способности молекул двухцепочечных РНК эффективно подавлять активность сходных с ними по строению генов. В последние годы РНК-интерференцию используют в прикладных исследованиях, направленных на получение нокаутных (содержащих «молчащие» гены) клеток, тканей и организмов. Ведь если грамотно использовать этот механизм, то теоретически можно «выключить» в клетке производство любого белка.
Среди успешных примеров применения этого метода — получение двух сортов кофейного дерева, содержащих в плодах пониженный на 30–50% алкалоид кофеина. Схожий эксперимент был проведен и с табаком с целью понизить в растении содержание никотина (Рябушкина, 2009).

Другая возможность использования этого подхода — подавление синтеза аллергенов. И это уже не сказка: генетикам из испанского Института сельского хозяйства в Кордове почти полностью удалось освободить зерна пшеницы от глиадина — одного из составляющих глютена. Именно из-за глиадина группа запасающих белков пшеницы вызывает у многих людей иммунную реакцию. Правда, и без использования системы редактирования генома CRISPR/Cas9 тут не обошлось (Sanchez-Leon et al., 2017).
Конечно, в этом направлении ученым еще работать и работать, но уже есть надежда, что совсем скоро можно будет смело есть арахисовое масло без боязни умереть от анафилактического шока! Важно отметить, что подобное вмешательство генной инженерии в метаболизм растений отличается от традиционного: в геном не встраивается чужеродный ген, а значит, не происходит и синтеза чужеродного белка. Из-за этого РНК-интерференцию можно назвать генным вмешательством со сниженным экологическим риском. Более того, даже формально такое вмешательство не несет на себе печати «ГМО».
Каков же итог нашей экскурсии по современным методам селекции? Шанс сделать выбор в пользу «натуральных» злаков, овощей и фруктов мы давно упустили. Прогресс, в том числе и в создании новых видов растений, остановить невозможно, но осознавать и правильно оценивать риски распространения и употребления генетически модифицированных продуктов необходимо.
Сегодня, когда общество проявляет большую озабоченность безопасностью пищевых продуктов, селекционеры находятся в ситуации, вынуждающей их использовать альтернативные пути получения новых сортов растений. В силу тех или иных причин эти методы не относятся к запрещенным, но в ряде случаев являются не менее рискованными, чем традиционные методики получения ГМО.
Также не стоит забывать, что грамотный подход к селекции растений с использованием техник редактирования генома позволяет минимизировать использование пестицидов и удобрений — что это означает для экологии, нет нужды объяснять. В любом случае, какие продукты мы будем есть завтра, в огромной степени зависит уже не от природы, а от нас самих.
Публикация подготовлена на основе статьи, которая участвовала в научно-популярном конкурсе «Био/мол/текст»-2017 портала «Биомолекула».
Литература
1. Закиян С. М., Власов В. В., Медведев С. П. «Редакторы геномов»: от «цинковых пальцев» до CRISPR // Наука из первых рук. 2014. Т. 56. № 2. С. 44–53.
2. Шумный В. К. Природа была первым генным инженером // Наука из первых рук. 2004. Т. 2. № 3. С. 32–39.
3. Cong L., Ran F. A., Cox D. et al. Multiplex Genome Engineering Using CRISPR/Cas Systems // Science. 2013. V. 339. P. 819–823.
4. From plant to crop: the past, present and future of plant breeding. 2016. VIB’s fact series, 44 p.
5. Philp J. C., Ritchie R. J., Allan J. E. Synthetic biology, the bioeconomy, and a societal quandary // Trends in Biotechnology. 2013. V. 31. P. 269–272.
6. Moghaddassi S., Eyestone W., Bishop C. E. TALEN-Mediated Modification of the Bovine Genome for Large-Scale Production of Human Serum Albumin // PLoS One. 2014, 9, e89631.
7. Matzke M. A., Kanno T., Matzke A. J. M. RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants // Annu. Rev. Plant Biol. 2015. V. 66. P. 243–267.
Трансформация генов или семь способов создать химеру
Время прочтения
9 мин
Просмотры 5.6K
Автор сообщества Фанерозой, биотехнолог, Людмила Хигерович.

Продолжаем знакомить вас с тонкостями работы биотехнологов. Готовы поспорить, что каждый из чтецов этого текста слышал о генно-модифицированных организмах, сокращенно ГМО. Кто-то их боится, кто-то считает их спасением от глобальных проблем человечества, кому-то абсолютно все равно.
Однако мы готовы поспорить, что большая часть читателей даже не представляет, как на практике создают этих самых ГМО или, как их правильнее называть в среде ученых, трансгенных организмов и генетических химер.
Справка
Химера или химерный организм — организм, несущий в себе генетический материал, принадлежащий двум и более видам живых существ. Химерой может быть как многоклеточный организм, так и одноклеточный. Кроме того, различают химер по тому, насколько и в каком качестве присутствует чужеродный геном: заимствована ли часть генов, встроены ли они в хромосому реципиента, или же оба генома сохранены полностью, но располагаются в разных клетках. Впрочем, это тема для отдельного поста.
Итак, чтобы создать генетически модифицированный организм, надо изменить его геном. В научной среде изменение генома называется трансформацией.
Трансформация генов – это изменение генетического состава клеток путем привнесения извне чужеродного генетического материала, перенос чужеродных (природных или искусственно созданных) донорских генов в клетки-реципиенты растений, животных и микроорганизмов, и получение таким образом трансгенных организмов с новыми или усиленными свойствами и признаками.
Секундное отступление. Трансформация — не изобретение человека. Бактерии умели передавать друг другу ДНК, захватывать чужую и встраивать ее в свой геном еще задолго даже до появления вирусов. Более того, эти свойства не утрачены даже у более развитых организмов. И, что еще более интересно, природная трансформация, также известная как горизонтальный перенос генов, играет огромную роль в эволюционных механизмах. Но это весьма обширная тема.
Стоит отметить, что трансформация имеет свои особенности для растений и животных. Растительные клетки из-за плотной клеточной стенки поверх мембраны неспособны самостоятельно поглощать фрагменты чужеродной ДНК и воспроизводить их, тогда как клетки животных могут различными способами поглощать фрагменты ДНК, встраивая их в свой геном, или поддерживать их воспроизведение в цитоплазме.
Исходя из этого учеными было разработано два основных метода генетической трансформации растений и еще пять – для животных. Разумеется, на самом деле их гораздо больше, однако по большей части это модификации основных семи. Рассмотрим вкратце каждый из них.
Векторный
По сути является лабораторной адаптацией механизма, называемого трансдукцией — обмен генами у бактерий и некоторых простейших. Пожалуй, он так же заслуживает отдельной статьи, но постараемся кратко.
Вектор — трансформирующий фактор, основой которого является самостоятельный фрагмент ДНК, несущий все необходимое для синтеза РНК и белка. Обычно на основе ДНК или РНК вируса, плазмиды или космиды (свободные генетические элементы бактерий и простейших).
В промышленной и сельскохозяйственной биотехнологии обычно создают векторы с помощью Ti-плазмид бактерий, так как они небольшие по объему, и относительно устойчивые.
Как их создают? Из плазмид выделяют участки Т-ДНК (транспортной), ответственные за встраивание в молекулу ДНК клетки-мишени и создают векторные ДНК (векторы), способные переносить и встраивать нужные последовательности, включая части, кодирующие нужный признак и вещество, а также маркеры (репортерные гены — они нужны для проверки — встроился ли вектор в ДНК реципиента).

Модифицированные агробактерии (или освобожденные плазмиды) помещают в одну среду с растительными клетками-реципиентами, откуда с помощью антибиотиков удаляют нетрансформированные клетки.
К векторной трансформации также относят перенос ДНК с помощью ретровирусов. Векторный перенос можно осуществить только при отсутствии клеточной стенки (т.е. необходимо привести клетку-донор в состояние протопласта).
Трансформированные клетки помещают в питательную среду для культивации (образование каллуса), впоследствии регенерируя из них целое растение.
- Плюсы: адресность (можно сделать так, чтобы вектор встраивался только в определенные участки), возможность проконтролировать встраивание вектора и его сохранность в следующих поколениях клеток.
- Минусы: большая подготовительная работа по синтезу и созданию вектора и его размножению, опасность случайных встраиваний.
Биобаллистический (Генный пистолет)
Биобаллистический или метод «генного пистолета» – применяют в основном в отношении однодольных растений, нечувствительных к агробактериям.
Пожалуй, это самый фантастический способ трансформации. И самый страшный для людей, которые всерьез боятся стать жертвами генной модификации. Ведь его принцип действия полностью соответствует его названию. Да, именно так — это пистолет, стреляющий… генами.

В специальных установках микрочастицы золота или вольфрама с нанесенной на них ДНК ускоряют при помощи сжатого гелия, и они проникают в ДНК клеток-мишеней. Как вариант этого метода встречается магнетофекция – перенос с помощью намагниченных металлических частиц.
- Плюсы: мобильность (можно таскать с собой в поле) и массовость (обработка большого количества клеток разом или целого растения).
- Минусы: внесенная ДНК практически ничем не защищена и может быстро потерять активность, нельзя проконтролировать встраивание.

Векторный и биобаллистический методы универсальны и для растений, и для бактерий, и для животных. Теперь же переходим к методам, характерным для трансформации животных клеток. В принципе, их так же можно применить на растительных клетках, однако придется лишить их клеточной стенки, и знатно повозиться с подготовкой клетки.
Метод микроинъекций
Это самый простой способ внести ДНК в клетку. Вы часто видели его в научно-фантастических фильмах на тему утечки какого-нибудь патогена из секретной лаборатории.
Суть ее заключается во введении целевых генов непосредственно в ядро животной клетки при помощи микрокапиллярной пипетки. Под микроскопом в ядро клетки вводится 1 × 10^(−10)—1 × 10^(−12) л раствора трансформирующей ДНК (несколько тысяч копий генов).

- Плюс: главный плюс этого метода в том, что введенный ген сохраняется внутри клетки, а клеточные структуры почти не повреждаются.
- Минус: при делении клетки только одна из дочерних клеток наследует дополнительную ДНК
Электропорация
Впервые была разработана в лаборатории Е. Неймана в 1982 т. для введения чужеродных нуклеиновых кислот в эукариотические клетки.
В многочисленных опытах было показано, что обработка клеток животных электрическими импульсами с напряженностью поля 5-10 кВ/см в течение 5-10 мкс приводит к поглощению клетками молекул ДНК, находящихся в среде.

- Плюсы: простота использования (кладем клетки в ванну с электролитом, капаем ДНКу, врубаем ток, профит!).
- Минусы: плохая контролируемость встраивания, повреждения ДНК и клеточных мембран.
Липофекция
С чем-то подобным сталкиваются блюстители красоты, пользующиеся лосьонами с мицеллами и принимающие жирорастворимые витамины.
Липосомы (мицеллы, везикулы) — “пузырьки” из одного или двух слоев липидов, по свойствам схожих с естественной клеточной мембраной. В основном различают так: липосомы — двухслойные, содержат комплекс гликолипидов, мицеллы — однослойные, преимущественно липидные. Но это не точно:)
Липосомный (везикулярный) метод – пиноцитозное (захват раствора) поглощение метафазных хромосом, заключённых в синтетическую фосфолипидную оболочку (линохромосом или липоплекса). Изолированные метафазные хромосомы способны проникать в чужеродную клетку и функционировать в ней в течение некоторого времени.

Большая часть поглощенных хромосом деградирует, распадаясь на отдельные фрагменты, осуществляющие перенос содержащихся в них генов. Фрагменты могут существовать в свободном состоянии. Гены трансгенома функционируют наряду с другими своими хромосомными генами клетки. Такие популяции клеток называют нестабильными.
При постоянном действии генов трансгенома появляются стабильные популяции клеток (клоны) – когда сохранившийся фрагмент интегрируется с хромосомой клетки реципиента. Интеграция происходит не путем рекомбинаций через двойной кроссинговер, а через транслоцирование фрагментов на негомологичные участки генома реципиента.

Соматическая гибридизация
Слияние соматических клеток с объединением их геномов — образование гибридома. Производится электрическим, химическим и механическим путем. Частный случай гибридизации – использование микрокеток, иногда объединяют с липофекцией.

Введение с помощью микроклеток – клетки доноров обрабатываются таким образом, что часть хромосом или отдельные хромосомы оказываются заключенными в часть цитоплазмы. Слияние микроклетки донора с полноценной клеткой реципиента ведет к тому, что реципиент получает группу или отдельные хромосомы донора.
Это один из самых трудоемких методов создания трансформированных клеток. Однако он полностью оправдывает себя — так, например, производят антитела для лечения редких заболеваний, иммуноглобулины и создают из клеток больных раком пациентов индивидуальные лекарства.
Химическая преципитация
Метод несколько схож с генным пистолетом с той лишь разницей, что частицы ДНК не выстреливаются, а доставляются на соли.
Например, преципитация фосфатом кальция выглядит следующим образом. ДНК добавляют к смеси раствора СаСl2 и фосфатного буфера, ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, 1973). Образуются частицы кальциевого преципитата. Они поглощаются клеткой путем фагоцитоза.
Основные методы трансформации закончились. Познакомимся с редкими, но все же встречающимися — некоторые из них весьма любопытны.
Органическая преципитация. Катионные полимеры
По сути, гибрид химической преципитации и фагоцитоза. В основном используют ДЕАЕ-декстран. Преципитация диэтиламиноэтил-декстраном (ДЕАЕ-декстраном) – использование положительно заряженных (катионных) водорастворимых полимеров, таких как ДЭАЭ-декстран, полиэтиленимин, поли-L-лизин или хитозан. Отрицательно заряженная ДНК связывает поликатионы, и образовавшийся положительно заряженный комплекс (полиплекс) взаимодействует с отрицательно заряженной мембраной клетки, что приводит к проникновению ДНК в клетку-мишень путем эндоцитоза. Метод считается малоэффективным из-за сложности с синтезом комплекса и слабом контроле доставки ДНК в ядро.

Фототрансфекция
Перенос фрагментов ДНК через поры, образованные в результате сверхкороткого облучения лазером. По сути, та же электропорация, только вместо тока — свет определенной длины волны.
Трансдукция
Вариация векторного метода для животных клеток. Имеет ряд отличий, например, в качестве вектора применяется фрагмент вирусной ДНК (ретровирусы, ДНК-вирусы, ВИЧ), чаще всего заключенный в реконструированную вирусную оболочку для облегчения проникновения в клетку и обеспечения тканеспецифичности.
Есть и еще менее популярные методы, как, например, трансформация тепловым шоком. Однако она сама по себе не привносит ДНК извне, лишь изменяя структуру существующей ДНК за счет температурных повреждений. Либо тепловой шок может служить тем же фактором повреждения мембраны, что и электрошок, открывая путь для проникновения ДНК. Но в таком случае, по сути он ничем не отличается от вышеназванных.
Итак, мы рассмотрели основные методы создания химерной (трансформированной) клетки или популяции клеток. Несмотря на то, что их характеристика по большей части уместилась в паре строк, каждый из них сопряжен с колоссальными усилиями как при подготовке, так и при исполнении, и тем более при контроле результата. Последнее особенно важно, так как любая клетка имеет свои механизмы защиты.
Мало трансформировать геном, надо еще обеспечить его сохранность и устойчивость — в противном случае, клетка быстро избавится от чужеродного фрагмента, вырезав его или растворив в лизосоме.
Для чего это делают?
На самом деле, трансформированные организмы давно уже нас окружают и служат нам. Стоит только подумать над тем, что первые научные работы на эту тему (на тему трансформации у бактерий) датированы до 1962 года, т.е. до подтверждения структуры ДНК! А сами бактериальные химеры стали применять аж в 1980-х.
Выше мы уже упоминали, что с помощью гибридов изготовляют лекарства от рака и редких заболеваний. Также с помощью трансформированных клеток производят гормоны (человеческий инсулин, ген которого перенесли в кишечную палочку), иммуноглобулины, белки, витамины (витамин С с помощью той же кишечной палочки, фитогормоны для растений с помощью сенной палочки), ферменты для пищевой, текстильной, химической промышленности, биоудобрения и биопрепараты для борьбы с паразитами и вредителями, антибиотики и даже очищают сахар (хотя это совсем уж редкий случай).
Вот вы узнали еще немного о биотехнологии.
Всего хорошего и не болейте!

Источники
Биотехнология: учебник и практикум для вузов / под редакцией Н. В. Загоскиной, Л. В. Назаренко. — 3-е изд., испр. и доп. — Москва: Издательство Юрайт, 2020. — 381 с.
Биотехнология / Т. Г. Волова. – Новосибирск: Изд-во Сибирского отделения Российской Академии наук, 1999. – 252 с
Биотехнология. Принципы и применение / Под ред. И.Хиггенса.- М.: Мир, 1980
Чечина, О. Н. Общая биотехнология: учебное пособие для вузов / О. Н. Чечина. — 2-е изд., перераб. и доп. — Москва: Издательство Юрайт, 2019. — 231 с.
Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998 Dec;16(6):735-43.
Godfrey J, Leukam MJ, Smith SM. An update in treating transformed lymphoma. Best Pract Res Clin Haematol. 2018 Sep;31(3):251-261.
Alton, E. W. F. W., Armstrong, D. K., Ashby, D., Bayfield, K. J., Bilton, Diana, Bloomfield, E. V.,… Wolstenholme-Hogg, P. (2015). Repeated nebulisation of non-viral CFTR gene therapy in patients with cystic fibrosis: A randomised, double-blind, placebo-controlled, phase 2b trial. Lancet Respiratory Medicine, 3(9), 684-691.
Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V., and Jackson, R. B. (2011). DNA tools and biotechnology. In Campbell biology (10th ed., pp. 408-435). San Francisco, CA: Pearson.
Transfection. Protocols and Applications Guide. Promega.
$begingroup$
Would I be able to genetically modify a plant at home? What equipment will be necessary? I think it might be a fun change from the ‘norm’ of regular hybridisation, to try some inter-family gene insertion, instead of staying within a genus. Are some plants easier to modify than others?
asked Jul 23, 2014 at 2:51
![]()
J. MusserJ. Musser
3,6776 gold badges37 silver badges74 bronze badges
$endgroup$
6
$begingroup$
Well, that depends on your home.  I think it is not an easy process.
I think it is not an easy process.
There are two main methods that are used to genetically modify plants:
Using the bacterium, Agrobacterium tumifaciens, as a vector for the DNA. Agrobacterium has the ability to infect plants and insert DNA into a plant’s genome. It causes crown gall tumours in natural infections. This method has mainly been used to modify broad leaved plants, such as sugar beet and oilseed rape, but is now also being applied to monocot species, such as maize and rice
Particle bombardment or biolistics where the DNA to be inserted is coated on minute gold particles and fired into plants cells. This approach is used for monocot plants such as maize and rice
Here I found simple step by step article, but lil bit old. may be there are new methods for this process.
How To Genetically Modify a Seed, Step By Step
answered Jul 23, 2014 at 4:26
![]()
I JI J
1,2261 gold badge10 silver badges15 bronze badges
$endgroup$
3
$begingroup$
Yes it is. The easiest plant to transform would be Arabidopsis, which can be transformed by agrobacterium using the floral dip method. The process would be as follows:
1. Design a gene sequence you wish to insert into the plant
2. Synthesize (or otherwise acquire the DNA)
3. Insert the DNA into your agrobacterium, at home you would use a cold snap transformation process described here: https://www.youtube.com/watch?v=PvkfIECvyqs This can be done at home using a dry ice bath.
4. Then use the floral dip method to insert the dna into the plant. This involves growing a plant to just the right age when it is flowering and dipping the Arabidopsis flowers into a solution of the agrobacterium.
5. Then grow the flowers to seed and grow the seeds on a suitable select able marker (which you designed into your DNA plasmid).
The specific experimental conditions needed to do all this are too long for this answer, but you can look them up online as all the procedures are standard.
Plants which result from this process would not be legal to release into the wild due to USDA regulations.
Engineering other plants, depending on the species, can be more complex but could possibly also done at home. There are even designs online for an open source gene gun if you didn’t want to use agrobacterium.
answered Sep 18, 2014 at 6:19
![]()
$endgroup$
$begingroup$
genetic modification can be done with mutations. A mutation is a permanent change in the sequence of DNA. In order to obtain an observable effect, mutations must occur in gene exons, or regulatory elements. Changes in the non-coding regions of DNA (introns and junk DNA) generally do not affect function.
Mutations can be caused by:
- external (exogenous) factors, such as chemicals and radiation
or
- endogenous (native) factors
Mutations can be advantageous and lead to an evolutionary advantage of a certain genotype..this will be a solution for your plant modification..
A mutagen is a physical or chemical agent that changes the genetic material, usually DNA, of an organism and thus increases the frequency of mutations above the natural background level.
chemical mutagens:
- Bromine
- Sodium azide
- Psoralen
- Benzene
long term treatment plant with mutagens caused mutations some might be advantage
then select advantage ..
As many mutations cause cancer, mutagens are therefore also likely to be carcinogens!
![]()
mgkrebbs
8,7892 gold badges28 silver badges46 bronze badges
answered Sep 20, 2014 at 17:50
![]()
$endgroup$
$begingroup$
answered Jul 23, 2014 at 4:24
![]()
Devashish DasDevashish Das
5,1823 gold badges22 silver badges68 bronze badges
$endgroup$
8
$begingroup$
Not to forget that simple cross breeding your plants the Mendelian way also modifies their genetic make up, as @souvik bhattacharya indicates in his comment. No equipment necessary, no chemicals, no ethical issues.
answered Nov 12, 2014 at 2:06
![]()
AliceD♦AliceD
51.8k18 gold badges169 silver badges236 bronze badges
$endgroup$
3
$begingroup$
Would I be able to genetically modify a plant at home? What equipment will be necessary? I think it might be a fun change from the ‘norm’ of regular hybridisation, to try some inter-family gene insertion, instead of staying within a genus. Are some plants easier to modify than others?
asked Jul 23, 2014 at 2:51
![]()
J. MusserJ. Musser
3,6776 gold badges37 silver badges74 bronze badges
$endgroup$
6
$begingroup$
Well, that depends on your home. I think it is not an easy process.
There are two main methods that are used to genetically modify plants:
Using the bacterium, Agrobacterium tumifaciens, as a vector for the DNA. Agrobacterium has the ability to infect plants and insert DNA into a plant’s genome. It causes crown gall tumours in natural infections. This method has mainly been used to modify broad leaved plants, such as sugar beet and oilseed rape, but is now also being applied to monocot species, such as maize and rice
Particle bombardment or biolistics where the DNA to be inserted is coated on minute gold particles and fired into plants cells. This approach is used for monocot plants such as maize and rice
Here I found simple step by step article, but lil bit old. may be there are new methods for this process.
How To Genetically Modify a Seed, Step By Step
answered Jul 23, 2014 at 4:26
![]()
I JI J
1,2261 gold badge10 silver badges15 bronze badges
$endgroup$
3
$begingroup$
Yes it is. The easiest plant to transform would be Arabidopsis, which can be transformed by agrobacterium using the floral dip method. The process would be as follows:
1. Design a gene sequence you wish to insert into the plant
2. Synthesize (or otherwise acquire the DNA)
3. Insert the DNA into your agrobacterium, at home you would use a cold snap transformation process described here: https://www.youtube.com/watch?v=PvkfIECvyqs This can be done at home using a dry ice bath.
4. Then use the floral dip method to insert the dna into the plant. This involves growing a plant to just the right age when it is flowering and dipping the Arabidopsis flowers into a solution of the agrobacterium.
5. Then grow the flowers to seed and grow the seeds on a suitable select able marker (which you designed into your DNA plasmid).
The specific experimental conditions needed to do all this are too long for this answer, but you can look them up online as all the procedures are standard.
Plants which result from this process would not be legal to release into the wild due to USDA regulations.
Engineering other plants, depending on the species, can be more complex but could possibly also done at home. There are even designs online for an open source gene gun if you didn’t want to use agrobacterium.
answered Sep 18, 2014 at 6:19
![]()
$endgroup$
$begingroup$
genetic modification can be done with mutations. A mutation is a permanent change in the sequence of DNA. In order to obtain an observable effect, mutations must occur in gene exons, or regulatory elements. Changes in the non-coding regions of DNA (introns and junk DNA) generally do not affect function.
Mutations can be caused by:
- external (exogenous) factors, such as chemicals and radiation
or
- endogenous (native) factors
Mutations can be advantageous and lead to an evolutionary advantage of a certain genotype..this will be a solution for your plant modification..
A mutagen is a physical or chemical agent that changes the genetic material, usually DNA, of an organism and thus increases the frequency of mutations above the natural background level.
chemical mutagens:
- Bromine
- Sodium azide
- Psoralen
- Benzene
long term treatment plant with mutagens caused mutations some might be advantage
then select advantage ..
As many mutations cause cancer, mutagens are therefore also likely to be carcinogens!
![]()
mgkrebbs
8,7892 gold badges28 silver badges46 bronze badges
answered Sep 20, 2014 at 17:50
![]()
$endgroup$
$begingroup$
answered Jul 23, 2014 at 4:24
![]()
Devashish DasDevashish Das
5,1823 gold badges22 silver badges68 bronze badges
$endgroup$
8
$begingroup$
Not to forget that simple cross breeding your plants the Mendelian way also modifies their genetic make up, as @souvik bhattacharya indicates in his comment. No equipment necessary, no chemicals, no ethical issues.
answered Nov 12, 2014 at 2:06
![]()
AliceD♦AliceD
51.8k18 gold badges169 silver badges236 bronze badges
$endgroup$
3
Статья на конкурс «био/мол/текст»: Испокон веков люди пытаются улучшить урожаи сельскохозяйственных культур. Статья повествует о том, какими средствами пользовались селекционеры на протяжении развития растениеводства, а особое внимание уделяется одному из самых современных и многообещающих методов молекулярной генетики растений — цисгеномике. Пора узнать, чем отличаются трансгенные организмы от цисгенных!
От неосознанного отбора к направленной селекции и генетической модификации растений
С незапамятных времен люди стремились улучшить качество и свойства используемых растений. Для этого они выбирали лучшие плоды и самые крупные зерна, бессознательно изменяя растения в нужном направлении. С развитием генетики, открывшей законы наследственности и изменчивости, появилась возможность осознанно управлять передачей необходимых признаков. Экспериментаторы поняли, что методом простого отбора человек не может получить принципиально новых свойств у разводимых организмов, так как при отборе можно выделить только те генотипы, которые уже существуют в популяции. Поэтому для получения новых сортов растений стали применять гибридизацию — скрещивание организмов с желаемыми признаками. Несмотря на то, что этот метод используется уже более века, процесс создания новых сортов остается очень трудоемким и требует многих лет напряженной работы. Современное сельское хозяйство испытывает острую необходимость в новых сортах, отличающихся устойчивостью к воздействию биотических и абиотических факторов окружающей среды, высокой продуктивностью и продолжительными сроками хранения урожая. На сегодняшний день показано, что такими качествами обладают дикие предки культивируемых растений. Чтобы передать их «полезные» гены современным сортам, необходимо проведение межвидового скрещивания, которое технологически сложнее и возможно далеко не для всех культур в силу генетической несовместимости.
Выход из этой ситуации появился с развитием генетической инженерии, которая сделала возможным перенос генов из одного организма в другой.
Различные виды генетически модифицированных растений
Генетически модифицированный организм (ГМО) — организм, генотип которого был целенаправленно изменен при помощи методов генной инженерии *. Основными способами биотехнологического изменения генома растений являются: искусственный мутагенез (физический и химический), трансгенез — введение гена неродственного организма, интрагенез — введение гена самогό организма или его «выключение», а также цисгенез — введение гена близкородственного вида, с которым возможно природное скрещивание.
* — На биомолекуле можно найти и размышления о пользе/вреде ГМ-растений, и «рецептуру приготовления» ГМ-риса и даже стратегии создания растительных фабрик: «Трансгенные растения — спасители планеты или бомбы замедленного действия?» [1], «Готовим ГМ-рис вместе» [2], «Растения-биофабрики» [3]. — Ред.
В 1994 году появилось первое коммерческое генно-инженерное растение — томат Flavr Savr [1, 4]. Вслед за этим стали активно создаваться трансгенные растения, устойчивые к биотическим и абиотическим факторам среды. (Отметим, что трансгенным считают такой организм, в геном которого искусственно введен ген, который не может быть приобретен при естественном скрещивании.) Ученые нашли необходимые гены устойчивости в геномах бактерий и насекомых и перенесли их в растительные организмы [5]. Обывателю покажется, что это достаточно рискованно, потому что, на первый взгляд, в природе такого быть не может (хотя есть и исключения — например, почвенная бактерия Agrobacterium tumefaciens переносит свои гены в растительные клетки для получения необходимых метаболитов).
Другая проблема, с которой связано получение трансгенных растений, — это использование антибиотиков. Для того чтобы узнать, попал ли интересующий ученых ген в геном растительной клетки, необходим некий маркер (репортер), который отделит клетки с внедрившимся чужеродным геном от неудачных образцов. Такими репортерами и являются гены, кодирующие устойчивость к антибиотикам. Клетки, подвергшиеся изменениям, высаживают на среду с антибиотиком, и, если они остались живы, значит, ген устойчивости проник в их геном, а с ним — и наш целевой ген. Несмотря на то, что трансгенные растения являются мощным фактором развития сельского хозяйства и экономики, возможность их использования провоцирует широкое общественное обсуждение.
Принимая во внимание всеобщую обеспокоенность биологической безопасностью трансгенных продуктов питания, в настоящее время активно разрабатывается новый подход для модификации сортов растений — цисгенез [5–7].
Применение геномного секвенирования сельскохозяйственно-значимых культур, таких как кукуруза, картофель, рис, и разработка эффективных технологий выделения новых генов расширили границы возможностей улучшения сельскохозяйственных культур. В последние десятилетия описан широкий круг генов, кодирующих важные качественные и количественные признаки как самих сельскохозяйственных культур, так и их дикорастущих родственников. Эти гены выделены и перенесены в геномы элитных сортов. Полученные в результате таких манипуляций растения называют цисгенными, чтобы отделить их от понятия трансгенов [5].
Цисгенез — такая технология генетической модификации рекомбинантной ДНК, при которой манипуляция происходит с использованием ДНК того же или близкородственного вида растения, с которым возможен половой процесс [8]. В отличие от трансгенных, такие растения не содержат гены неродственных организмов и гены устойчивости к антибиотикам (рис. 1). Это дает возможность ожидать, что общество с большей легкостью воспримет цисгенные растения, нежели трансгенные. Так, опрос в штате Миссисипи показал, что 81% респондентов готов употреблять в пищу цисгенные растения, в то время как лишь 14-23% согласны на трансгенные [5].
| Трансгенез | Цисгенез | Интрагенез |
|---|---|---|
|
|
|

Рисунок 1. Изменение генома при классической селекции, трансгенезе и цисгенезе.
Цисгенез = классическая селекция?
Современное состояние вопроса о повышении качества и урожайности сельскохозяйственных растений можно объяснить на примере такой стратегически важной культуры, как картофель.
Представим себе, что у нас есть элитный сорт картофеля, который дает прекрасный — качественный и обильный — урожай, но подвержен инфекционным заболеваниям. А еще нам известно, что есть вид дикого картофеля, который не дает никакого съедобного урожая, но при этом устойчив к болезням. Первым делом мы пытаемся выяснить, что в геноме дикого вида определяет его устойчивость (см. врезку). Возможности современной молекулярной генетики и геномики растений позволяют нам найти тот ген, который отвечает за устойчивость, выделить его открытую рамку считывания и участок ДНК, контролирующий его экспрессию, вырезать их, клонировать и внедрить в геном нашего элитного сорта, не внося никаких кардинальных изменений ни в геном сорта, ни в нуклеотидную последовательность гена, которой мы оперируем. Таким образом, если наши манипуляции прошли успешно, мы получаем исходный элитный сорт картофеля, который так же дает прекрасные клубни, но при этом не подвержен заражению паразитами, и мы не теряем урожай.
На сегодняшний день цисгеномика является альтернативным подходом, а основным инструментом создания новых сортов растений остаются классические методы селекции. Для этого мы скрещиваем наш элитный сорт с диким видом картофеля, получаем гибрид, у которого половина генетического материала происходит от исходного элитного сорта, а половина — от дикого предка. Таким образом мы приобретаем устойчивый к паразитам гибрид. Заметим, что мы при этом теряем половину полезных генов, которые были у элитного сорта, и наш новый гибрид не дает такого обильного и вкусного урожая, который долго хранится и не боится холодов. Более того, помимо желаемого гена устойчивости от дикого предка наш гибрид получает еще тысячи ненужных (а возможно, и опасных) сцепленных с ним генов, продукты которых могут оказаться токсичными. Подобная ситуация нас не устраивает, и мы проводим так называемое «обратное насыщающее скрещивание»: пытаемся вернуть нашему гибриду «утерянные» гены элитной родительской формы. Для этого мы опять скрещиваем наш гибрид с элитным сортом, тем самым «разбавляя» его генόм. И так множество раз. Заметим, что после каждого скрещивания должно пройти достаточно времени, чтобы новый гибрид вырос и дал урожай. В результате процесс классической селекции обычно затягивается на десятки лет.
Если вы всё еще против генетически модифицированных растений, представим следующую ситуацию: у нас есть элитный сорт картофеля, единственным слабым местом которого является подверженность заболеваниям. Классическая селекция отнимает слишком много сил и времени и не дает надежного результата, а генетически модифицированные растения не внушают доверия. Даже в этой ситуации есть альтернатива! Химическая промышленность изобрела огромное количество разнообразных пестицидов, которые защитят наш картофель от болезней. Этот путь самый простой и самый опасный: загрязнение окружающей среды химическими реагентами может привести к катастрофическим последствиям (см. врезку).
| Метод | Принцип метода | Достоинства | Недостатки | |
|---|---|---|---|---|
| Классическая селекция | Гибридизация | Многократные скрещивания видов или сортов с интересующими селекционера признаками с дальнейшим искусственным отбором | Проверено временем, не вызывает опасений в обществе | Требует больших временных затрат (от трех до 15 лет) Скрещивание возможно не для всех растений |
| Искусственный мутагенез | Физическая (радиация) и химическая (супермутагены) обработка семян растений и самих растений с дальнейшим жестким отбором | Образование новых признаков и свойств | Требует больших временных затрат Низкая продуктивность, опасность для здоровья экспериментатора |
|
| ГМО | Трансгенез | Ген, определяющий хозяйственно важные признаки, переносится в геном изменяемого растения из бактерии, насекомого или другого неродственного организма | Существует большое разнообразие потенциально полезных генов, которые можно перенести в культурные растения | Вызывает опасения в обществе |
| Интрагенез | Улучшение свойств растения за счет внедрения в его геном дополнительной копии своего же гена (сверхэкспрессия) | Не используются чужеродные гены | Мало потенциально значимых генов в геноме самого растения, при этом многие признаки могут иметь полигенную природу | |
| Цисгенез | В геном элитного сорта переносится ген из близкородственного вида растения, с которым потенциально возможно скрещивание в природе | Результат метода абсолютно аналогичен классической селекционной работе, но при этом существенно экономит время и гарантирует отсутствие попадания в геном нежелательных сцепленных генов растения-донора | Круг интересных хозяйственно важных генов меньше по сравнению с трансгенными растениями | |
| Другие способы повышения продуктивности растений | Химические реагенты | Химические реагенты распыляются в большом количестве на поля и напрямую убивают вредителей растений (бактерий, грибы, сорняки и насекомых), тем самым повышая урожайность | Скорость | Наносится существенный вред окружающей среде, нарушается баланс в природе. Химические вещества накапливаются в растениях и продуктах питания, в результате попадая в организм потребителей |
| Биологические способы | Обработка тепличных растений
|
Безопасность | Высокая стоимость |
Итак, цисгенные растения, главной целью создания которых является перенесение генов устойчивости в коммерчески успешные сорта, экономят время селекционеров, не требуют применения пестицидов, не нарушают экосистему, затраты на их выращивание минимальны, а урожай максимальный. В 2012 году Европейское агентство по безопасности продуктов питания (EFSA) опубликовало доклад, в котором сравнивался потенциальный вред растительных продуктов, полученных различными способами. В результате был сделан вывод о том, что риски, связанные с употреблением цисгенных растений и сортов, полученных методами классической селекции, сопоставимы [17].
Однако формально цисгенные растения подходят под определение генетически модифицированных организмов. В России правовое регулирование данной области базируется на Федеральном законе от 05.07.1996 № 86-ФЗ «О государственном регулировании в области генно-инженерной деятельности». В 2015 году правительство приняло законопроект об отказе от применения технологий генетической модификации при производстве продуктов питания. Если ситуация изменится, использование этой технологии приведет к бурному развитию сельского хозяйства.
- Трансгенные растения — спасители планеты или бомбы замедленного действия?;
- Готовим ГМ-рис вместе;
- Растения-биофабрики;
- Википедия: «Flavr Savr» (англ.);
- Telem R.S., Wani S.H., Singh N.B., Nandini R., Sadhukhan R., Bhattacharya S., Mandal N. (2013). Cisgenics — a sustainable approach for crop improvement. Curr. Genomics. 14, 468–476;
- Jacobsen E. and Schouten H.J. (2008). Cisgenesis, a new tool for traditional plant breeding, should be exempted from the regulation on genetically modified organisms in a step by step approach. Potato Research. 51, 75–88;
- Espinoza C., Schlechter R., Herrera D., Torres E., Serrano A., Medina C., Arce-Johnson P. (2013). Cisgenesis and intragenesis: new tools for improving crops. Biol. Res. 46, 323–331;
- Schouten H.J., Krens F.A., Jacobsen E. (2006). Cisgenic plants are similar to traditionally bred plants: international regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep. 7, 750–753;
- Ellis J. and Jones D. (1998). Structure and function of proteins controlling strain-specific pathogen resistance in plants. Curr. Opin. Plant Biol. 1, 288–293;
- Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов;
- Flor H.H. (1971). Current status of the gene-for — gene concept. Ann. Rev. Phytopathol. 9, 275–296;
- Шамрай С.Н. (2003). Гены устойчивости растений: молекулярная и генетическая организация, функции и эволюция. Журнал общей биологии. 64, 195–214;
- Чекалин Н.М. Генетические основы селекции зернобобовых культур на устойчивость к патогенам. Полтава: Iнтерграфiка, 2003. — 186 с.;
- Шорохов М.Н. Рынок средств защиты растений (СЗР) в мире и России. СПб: Санкт-Петербургский государственный аграрный университет, 2003. — 25 с.;
- Куценко С.А. Основы токсикологии. М.: Фолиант, 2004. — 570 с.;
- Википедия: «Бхопальская катастрофа»;
- Andersson H.C., Arpaia S., Bartsch D., Casacuberta J., Davies H., du Jardin P. et al. (2012). Scientific opinion addressing the safety assessment of plants developed through cisgenesis and intragenesis. EFSA Journal. 10, 2561..
ГЕННАЯ ИНЖЕНЕРИЯ РАСТЕНИЙ
5.1 Проблемы биобезопасности
5.2. Получение трансгенных растений
5.3. Применение методов генетической инженерии для улучшения аминокислотного состава запасных белков растений
5.4. Повышение эффективности процесса фотосинтеза
5.5. Генно-инженерные подходы к решению проблемы усвоения азота
5.6. Изменение качества плодов
5.7. Устойчивость растений к гербицидам
5.8. Устойчивость растений к фитопатогенам и насекомым
5.9. Устойчивость растений к абиотическим стрессам
Генно-инженерные методы, в частности технология рекомбинантных ДНК, позволяют создавать новые генотипы и, следовательно, новые формы растений гораздо быстрее, чем классические методы селекции. Кроме того, появляется возможность целенаправленного изменения генотипа — трансформации — благодаря введению определенных генов.
Считается, что трансгенные растения более безопасны, чем сорта, полученные методами традиционной селекции, но с применением мутагенеза (В. Ф. Федоренко и др., 2005). Есть мнение, что риск использования трансгенных растений в мирных биотехнологиях намного ниже, чем в случае применения других трансгенных организмов (Я. И. Бурьянов, 1999). Генетическая инженерия растений развивается очень быстрыми темпами. Она позволяет решать проблемы фундаментальной науки, а также многочисленные практические задачи. Намечается тенденция разделения прикладных генно-инженерных исследований на три направления:
- Решение проблем сельскохозяйственного комплекса, таких, как повышение продуктивности сельскохозяйственных растений, их защита от различных биотических и абиотических факторов. Это особенно важно в связи с быстрым ростом населения Земли. Считается, что генетически модифицированные (ГМ) растения помогут решить проблему увеличения производства продуктов питания, поэтому сейчас наблюдается весьма значительное расширение сельскохозяйственных угодий, занятых посевами ГМ — растений. Если в 1996 г. эти посевы занимали 1,7 млн га, то в 2001 г. — 52,6 млн га, а в 2003 г. — почти 70 млн га.
- Создание трансгенных растений — продуцентов новых веществ для использования в области медицины и технической промышленности, приспособленных для роста в полевых условиях. Так, на основе трансгенных растений возможно получение «съедобных» вакцин, антител (например, иммуноглобулиновых комплексов).
Развитие и совершенствование культуры трансгенных клеток и тканей растений, синтезирующих ценные биологически активные соединения. Довольно часто продуцентами важных пищевых, лекарственных или технически важных веществ служат уникальные тропические и эндемические растения, не доступные для выращивания в других климатических зонах или занесенные в «Красную книгу». Перенос генов, отвечающих за синтез специфических соединений, из редких растений в более доступные превращает последние в биологические фабрики необходимых веществ.
Генетическая трансформация может принести большую пользу для сельского хозяйства, медицины, промышленности, фундаментальной науки.
Однако биотехнология и, в частности, генетическая инженерия подошли к той ступени развития, когда приходится прежде всего думать о возможных последствиях эксперимента, об использовании полученных знаний. «Генетическая инженерия — это мощный способ изменить жизнь, но ее потенциал может представлять собой опасность» (Р. Доукинс, цит.: Современная биотехнология. Мифы и реальность, 2004). В связи с этим первостепенными становятся вопросы биологической безопасности.
Под биологической безопасностью понимается защищенность человека, общества, цивилизации и окружающей среды от вредного воздействия, опасного для жизни и здоровья людей, от токсичных и аллергенных биологических веществ и соединений, содержащихся в природных или генетически модифицированных биологических объектах и полученных из них продуктах (О. С. Машкина, А. К. Буторина, 2005). В настоящее время выделяют три группы риска, возникающие при возделывании и потреблении генетически модифицированных организмов (ГМО): пищевые, экологические, агротехнические. Причины возникновения этих рисков состоят в непредсказуемости встраивания дополнительной ДНК в геном организма, возможности плейотропного эффекта встроенного гена, нарушении стабильности генома трансгенных растений, присутствии во встраиваемой конструкции «лишних» векторных генов, аллергических и токсических эффектах чужеродного белка, возможности горизонтального переноса трансгенных конструкций в геном симбионтных для человека и животных бактерий (В. В. Кузнецов и др., 2004). По-видимому, есть закономерность между увеличением числа аллергических заболеваний и потреблением ГМ-про- дуктов в странах, где это потребление разрешено. Так, в США и Скандинавии был проведен сравнительный анализ количества пищевых заболеваний. Эти страны были выбраны в связи с тем, что единственным существенным различием в качестве питания были потребление генетически модифицированных продуктов в США и их отсутствие в рационе народов Скандинавии. Оказалось, что в США частота пищевых заболеваний в 3 — 5 раз выше
чем в странах Скандинавии (В. В. Кузнецов и др., 2004). Нельзя исключить, что генетическая инженерия может привести к результатам, опасным для человека и природы. Поэтому несомненно, что биологическая безопасность должна стать одной из приоритетных задач человечества в целом и каждой цивилизованной страны в отдельности (А.С.Спирин, 1997, цит.: В.В.Кузнецов и др., 2004).
5.2. Получение трансгенных растений
Технологии получения трансгенных растений практически совпадают с теми технологиями, которые обсуждались в учебном пособии, когда речь шла о передаче генетической информации у микроорганизмов и животных. Поэтому в данном разделе рассмотрим только те системы, которые специфичны для растительных организмов.
Векторы на основе Ti-плазмид. Перенос генов и их встраивание в геном двудольных растений (трансформация) осуществляются главным образом благодаря конструированию векторов, характерных для растений. Эти векторы создаются на основе Ti-плазмид, находящихся в клетках бактерий Agrobacterium faciens.
Среди многочисленных почвенных бактерий существуют бактерии рода Agrobacteria, которые могут передавать часть своих генов в геном растительной клетки, вызывая коренные изменения в ее свойствах. Так, бактерия A. tumefaciens вызывает образование опухолей — корончатых галлов. Способность этой бактерии к образованию опухоли связана с геномом большой внехромосомной плазмиды —Ti-плазмиды (от англ. tumor inducing — индуцирующие опухоль). Другие бактерии ( . rhizogenes) вызывают усиленное образование корней при заражении растений. За этот процесс ответственны содержащиеся в бактериальных клетках Ri- плазмиды (от англ. root inducing — индуцирующий корни). Ti- и Ri-плазмиды — естественные векторы для дополнительных генов, обладающие свойствами, которые необходимы для трансформации растительных клеток. Они имеют широкий круг хозяев, а в бактериальных клетках реплицируются автономно. Ri-плазмиды выгодно отличаются от Ti-плазмид тем, что они не онкогенны. Клетки растений, трансформированные с помощью Ri-плазмид, сохраняют способность к морфогенезу и регенерации растений. В связи с этим Ri-плазмиды в данный момент рассматриваются как более перспективные для создания векторов. Однако Ti-плазмиды изучены лучше, поэтому их, как правило, используют при работе с растениями.
Доказательством того, что именно Ti-плазмиды отвечают за трансформацию растительных клеток в опухолевые, служит тот факт, что клетки агробактерий, лишенные Ti-плазмид, не могут вызвать в инфицированном растении ни образования корончатых галлов, ни синтеза опинов. Опины — это уникальные продукты конденсации амино- и кетокислот или аминокислот и сахаров, которые используются бактериями в качестве источников азота, углерода и энергии.
После заражения часть Ti-плазмиды встречается в хромосомах клеток растения-хозяина. Следовательно, A. tumefaciens встраивает часть своего генома в ДНК растительной клетки и заставляет ее таким способом изменять метаболизм, синтезируя вещества, необходимые для бактерий. Сама бактерия в растительную клетку не проникает. Она остается в межклеточном пространстве, используя трансформированные клетки растений в качестве биологической «фабрики» по производству опинов.
Участок Ti-плазмиды, который встраивается в геном растительных клеток, называется Т-ДНК — трансформирующая ДНК (transferred DNA). Величина Т-ДНК составляет примерно 10% от размера всей Ti-плазмиды, т.е. 12—22 тыс. п.н. Т-ДНК содержит гены, отвечающие за индукцию образования опухоли, синтез опинов и фитогормонов (ауксинов и цитокининов), подавляющих дифференцировку клеток растения-хозяина. Гены, отвечающие за вырезание Т-ДНК и ее перенос в растительную клетку, расположены в области вирулентности (vir-область) той же самой плазмиды. Кроме того, в состав Ti-плазмиды входят гены tra-области, контролирующие конъюгацию бактерий, и гены ori-области, продукты которых обеспечивают репликацию Ti-плазмиды (рис. 5.17).
Молекулярно-генетический механизм естественной трансформации растительных клеток с помощью Ti-плазмид изучен достаточно хорошо. В этом процессе можно выделить несколько этапов:
- восприятие раневых си гнал ор растительной клетки хеморецепторами бактерии и прикрепление агробактерии к стенке клетки растения;
- вырезание Т-ДНК из Ti-плазмиды;
- проникновение Т-ДНК внутрь клетки растения;
- интеграция Т-ДНК в геном растения и ее экспрессия.
Трансформация начинается только там, где есть поврежденные клетки. Весь процесс «узнавания» нужной растительной клетки, вырезания, транспорта и встраивания Т-ДНК в растительный геном осуществляют белки — продукты генов, локализованных в vir-области Ti-плазмиды. Продукты гидролиза поврежденной клеточной стенки (ацетосирингон и гидроксиацетосирингон) воспринимают хеморецепторы, встроенные в плазматическую мембрану агробактерии.

Среди них одна из главных ролей принадлежит продукту деятельности генов vir-области — гистидиновой протеинкиназе, которая активирует транскрипцию остальных vir- генов. Индукция этих генов обратима, что очень важно для бактерий. Если растительный организм ослаблен или вообще нежизнеспособен, процесс останавливается и Т-ДНК не переносится в такую клетку. Другой продукт генов vir-области — эндонуклеаза, которая отвечает за вырезание Т-ДНК. Белки virB и virE транспортируют Т-ДНК в одноцепочечной форме из бактерии в цитоплазму растительной клетки. Механизм переноса Т-ДНК, по-видимому, аналогичен переносу плазмидной ДНК в процессе конъюгации. Встраивание Т-ДНК в геном клетки растения становится возможным благодаря наличию гомологичных участков между растительной и плазмидной ДНК в местах встраивания. После внедрения Т-ДНК становится обычной частью генома растения и наследуется как доминантный признак в соответствии с законами Менделя (рис. 5.18).

Именно способность участка Ti-плазмиды A. tumefaciens встраиваться и экспрессироваться в геноме растительной клетки послужила поводом для попытки создания на основе этой плазмиды вектора, доставляющего дополнительные гены в клетку. Т-ДНК ограничена с двух сторон повторяющимися последовательностями (фланкирующими последовательностями), по которым и происходит вырезание этого участка ДНК из плазмиды. Казалось бы, что любая ДНК, встроенная между этими последовательностями, будет принята за Т-ДНК и перенесена в растительную клетку любых двудольных растений.
Однако,Ti-плазмида как вектор обладает некоторыми недостатками: большие размеры Ti-плазмиды (от 200 до 800 тыс. п. н.); отсутствие способности реплицироваться в клетках Е. coli, что необходимо для клонирования рекомбинантных плазмид; неспособность трансформированных с помощью Ti-плазмид клеток к регенерации целого растения; неспособность к трансформации клеток почти всех однодольных растений. Основной недостаток — слишком большие размеры Ti-плазмиды, которые делают весьма затруднительными манипуляции с ней во время конструирования вектора. Для решения этой проблемы разработана технология создания промежуточного вектора с применением классического объекта генетической инженерии – Е.coli.
Процесс начинается с создания рекоминантной структутры, которую можно достаточно просто клонировать, т.е. многократно увеличивать число ее копий. Т-ДНК вырезают в помощью рестриктаз из Ti-плазмиды и встраивают в небольшую плазмиду pBR322, способную реплицироваться в клетках E.coli. В этих клетках рекомбинантную ДНК клонируют, после чего встраивают в Т-ДНК дополнительные гены и повторяют клонирование. Полученные многочисленные копии промежуточной векторной ДНК вводят в клетки A. tumefaciens, содержащие обычные Ti- плазмиды. В результате гомологичной рекомбинации Т-ДНК с встроенными в нее дополнительными генами замещает нормальную Т-ДНК в Ti-плазмидах. После этого отбирают с помощью генов-маркеров трансформированные агробактерии и заражают ими растительные клетки. Дополнительные гены вместе с Т-ДНК встраиваются в геном растения, и в конце концов образуется генетически модифицированный организм с заданными свойствами (рис. 5.19). Эту довольно сложную технологию можно упростить, если использовать бинарные векторы.

Бинарные векторы представляют собой бактерии, содержащие две разные Ti-плазмиды. Одна из них несет vir-область и обеспечивает интеграцию в геном растительной клетки Т-области, содержащей любые гены другой плазмиды. В этом случае двойной кроссинговер не требуется.
Векторы на основе ДНК-содержащих вирусов растений. Вирусы можно рассматривать как разновидности чужеродной нуклеиновой кислоты, которые реплицируются и экспрессируются в клетках растений. Подавляющее большинство фитовирусов в качестве носителя генетической информации содержат РНК. Только 1 — 2% вирусов, инфицирующих растения, относятся к ДНК-содержащим. Именно эти вирусы удобны для использования в технологии рекомбинантных ДНК, а также в качестве векторов.
ДНК-содержащие вирусы могут включать одноцепочечную или двухцепочечную ДНК. В качестве представителей первой группы можно назвать вирус золотистой мозаики фасоли (ВЗМФ) или вирус полосатости кукурузы. Наиболее изученный представитель группы вирусов с двухцепочечной ДНК — вирус мозаики цветной капусты (ВМЦК), поражающий в основном растения семейства крестоцветные.
Обычно фитовирусы реплицируются с образованием большого числа копий молекул нуклеиновых кислот — 10 и более молекул на зараженную клетку. Поэтому фитовирусы представляют собой очень эффективные средства для получения хорошей экспрессии чужеродного гена. Кроме высокой копийности вирусной нуклеиновой кислоты вирусные векторные системы имеют еще ряд преимуществ: малый размер генома (возможность легкой мани пуляции вирусной ДНК) и сильные промоторы, обеспечивающие эффективную экспрессию чужеродных генов.
Однако вирусы в качестве векторов обладают и существенными недостатками: имеют небольшую емкость, патогенны и неспособны встраиваться в хромосомы хозяина. Небольшую емкость можно увеличить, если инфицировать вирусом (например, ВМЦК) растительные протопласты, а не клетки. В этом случае инфекция не передается от клетки к клетке, нет необходимости в упаковке ДНК в вирусные частицы. Следовательно, часть вирусного генома, ответственная за упаковку в вирусные частицы, может быть удалена и замещена дополнительной чужеродной ДНК. Другой недостаток — отсутствие способности встраиваться в геном растительной клетки — удается обойти (по крайней мере для ВМЦК) благодаря специальному методическому приему — агроинфекции. Для этого геном ВМЦК встраивают в Т-область Ti- плазмиды и в ее составе интегрируют в ядерный геном различных растений.
Методы прямого переноса генов в растение. Эти методы стали использоваться для растений благодаря появлению специфического объекта — изолированных протопластов, т. е. клеток, лишенных целлюлозной стенки.
Методы прямого переноса генов довольно многочисленны:
Трансформация растительных протопластов. Метод осуществляется благодаря комбинации методик кальциевой преципитации ДНК и слияния протопластов. Для трансформации может быть использован практически любой ДНК-вектор. Донорная ДНК может не содержать специальных биологических сигналов (vir-областей, пограничных областей Т-ДНК).
Заражение культуры протопластов на начальной стадии ее роста агробактериями, которые используют в качестве векторов.
Микроинъекции ДНК. Этот метод аналогичен методу микроинъекций животных клеток. Его можно рассматривать как наиболее универсальный. Эффективность трансформации растительных клеток — 10 — 20 % независимо от типа вектора. Трансформация не видоспецифична, возможен перенос генов в любое растение.
Электропорация. Метод основан на повышении проницаемости биомембран за счет действия импульсов высокого напряжения. В результате молекулы ДНК проникают в клетки через поры в клеточной мембране.
Упаковка в липосомы. Это один из методов, позволяющих защитить экзогенный генетический материал от разрушения нуклеазами растительной клетки. Липосомы — сферические тельца, оболочки которых образованы фосфолипидами.
Метод биологической баллистики. Это один из самых эффективных методов трансформации однодольных растений. Исходный материал для трансформации — суспензионная культура, каллусная ткань или 4 — 5-дневные культивируемые незрелые зародыши однодольных.
Метод основан на напылении ДНК-вектора на мельчайшие частички вольфрама, которыми затем бомбардируют клетки. Бомбардировка осуществляется с помощью биолистической пушки за счет перепада давления. Часть клеток гибнет, а выжившие клетки трансформируются, затем их культивируют и используют для регенерации растений.
5.3 Применение методов генетической инженерии для улучшения аминокислотного состава запасных белков растений
Решение проблемы создания новых форм растений подразумевает в первую очередь повышение качества синтезируемых растением продуктов, которые определяют его питательную и техническую ценность. В основном это касается запасных белков.
В большинстве случаев запасные белки растений имеют несбалансированный для питания человека и животных аминокислотный состав. Так, запасные белки злаков — проламины — бедны лизином, триптофаном и треонином, что снижает их питательную и кормовую ценность. Улучшение аминокислотного состава белка путем традиционной селекции не дает желательных результатов, поскольку необходимые гены часто сцеплены с нежелательными признаками и наследуются вместе. Например, у мутантов кукурузы и ячменя повышение содержания лизина коррелировало с уменьшением синтеза основных запасных белков — зеина и гордеина, а также с уменьшением урожайности.
Операции по получению трансгенных растений с улучшенным аминокислотным составом белка разделены на ряд этапов:
1) клонирование генов запасных белков;
2) изучение механизмов тканеспецифичной и временной экспрессии белков и выявление последовательностей ДНК, определяющих данный механизм;
3) целенаправленное изменение последовательностей генов запасных белков для улучшения аминокислотного состава;
4) создание векторов, содержащих измененный ген;
5) введение модифицированных генов в растения.
В настоящее время клонированы 10 генов гордеинов ячменя, гены а- и б-глиадинов и глютенина пшеницы, зеинов кукурузы, легумина бобовых, пататина картофеля и ряд других. Имеются практические результаты трансформации растений. Так, введение в геном пшеницы модифицированного гена проламина привело к активному синтезу модифицированного белка, а также повлияло на состав и уровень соответствующих запасных белков. В итоге улучшилось хлебопекарное качество пшеничной муки.
5.4. Повышение эффективности процесса фотосинтеза
Один из возможных способов увеличения фотосинтеза и, следовательно, продуктивности растений состоит в клонировании хлоропластных генов в клетках бактерий и их переносе в растения. Известно, что хлоропласты и прокариотические клетки сходны по ряду признаков. На основании этого возникла симбиотическая гипотеза происхождения хлоропластов, впервые выдвинутая А. С. Фаминциным (1886). Согласно этой гипотезе, клетки прокариот и хлоропласты сходны. В них присутствуют кольцевые ДНК, 708-рибосомы; синтез белков начинается с одной и той же аминокислоты — N-формилметионина, а синтез белка подавляется хлорамфениколом, а не циклогексимидом, как у эукариот. Позже было показано, что ДНК-зависимая РНК- полимераза Е. coli связывается с определенными участками ДНК хлоропластов шпината.
В клетках Е. coli инициация белкового синтеза частично регулируется доступностью участка связывания рибосом (УСР). Его структура до конца еще не выяснена, но расшифрован участок, известный под названием «последовательность Шайн—Дальгарно» (ШД). Она комплементарна б-концу 168-рибосомальной РНК, и инициация белкового синтеза начинается с образования комплементарной пары между этой последовательностью и 3′-концом 168-рибосомальной РНК. Анализ последовательности ДНК хлоропластного гена большой субъединицы основного фермента фотосинтеза — рибулозобисфосфаткарбоксилазы — оксигеназы (Рубиско) кукурузы — выявил значительные гомологии с известными промоторами и последовательностями ШД клеток Е. coli. Все это привело к попытке клонирования генов хлоропластов в клетках Е. coli, наиболее часто используемой в генно-инженерных исследованиях.
Транскрипционные конструкции могут создаваться двумя путями: во-первых, это установка промотора рядом с УСР, который узнается полимеразой Е. coli, во-вторых, это формирование гибридного УСР, состоящего из прокариотической последовательности ШД и эукариотического или синтетического инициирующего кодона. Первый путь широко применяется для увеличения экспрессии генов Е. coli и хлоропластных генов в клетках Е. coli.
В настоящее время уже клонировано несколько хлоропластных генов: гены синтеза субъединиц Рубиско, белка хлорофилл-белкового комплекса, АТФсинтетазы, цитохрома и др.
5.5. Генно-инженерные подходы к решению проблемы усвоения азота
Азот — один из самых необходимых элементов для растений. Его недостаток в почве или питательном субстрате часто приводит растение к гибели, поэтому в первую очередь необходимо внесение в почву азотных удобрений. Однако их производство требует очень больших энергетических затрат, поэтому оно дорогостояще. Стоимость азотных удобрений в 6 раз выше стоимости фосфорных удобрений и в 16 раз выше стоимости калийных удобрений. При этом растения используют только от 30 до 70 % внесенных в почву доступных форм азота, остальное просто вымывается из почвы, загрязняя окружающую среду. Гораздо более естественно и доступно снабжение растений азотом путем его биологической фиксации.
Фиксация атмосферного азота (диазотрофность) — свойство прокариотических организмов. Азотфиксирующие организмы делятся на симбиотические (90 %) и свободноживущие (10 %). Фиксация атмосферного азота связана преимущественно с симбиотическими микроорганизмами. В настоящее время известны четыре основные системы симбиоза, имеющие большое значение не только для естественных сообществ, но и для сельского хозяйства, лесоводства. Это Rhizobia — бобовые растения, Azolla-Anabaena — рис, Actinomyces — деревья, Spirillum — травы. Атмосферный азот фиксируется благодаря уникальному ферменту — нитрогеназе.
В 1960 г. американские исследователи показали, что нитрогеназа сохраняет свою активность в бесклеточных экстрактах Clostridium pasteurianum. Это послужило толчком для начала активных исследований биохимии азотфиксации, структуры и механизма действия нитрогеназы. К 1981 г. нитрогеназа была выделена из 36 видов микроорганизмов. Она считается одним из наиболее сложных ферментов, использующих простые субстраты. Кроме азота нитрогеназа может восстанавливать ацетилен, цианистый водород, закись азота и некоторые другие соединения. Восстановление ацетилена в этилен позволило разработать надежный тест для обнаружения азотфиксирующей активности. Непременное условие работы нитрогеназы — ее защита от кислорода, который ингибирует не только активность нитрогеназы, но и ее биосинтез.
Начиная с 1970 г. стали появляться серьезные работы по изучению генов азотфиксации и их переносу в клетки Klebsiella pneumoniae и Е. coli. С помощью техники рекомбинантных ДНК были составлены генетические карты генов азотфиксации (nif-генов), которые показали сходную организацию генов у большей части азотфиксирующих организмов. Было установлено, что nif- гены расположены между генами, кодирующими биосинтез гистидина (his) и генами, ответственными за усвоение шикимовой кислоты (shiA). Гены, кодирующие синтез белковых субъединиц компонентов нитрогеназы, образуют единый оперон (рис. 5.20). В клетках симбиотических бактерий Rhizobium leguminosarum, R. meliloti, R. trifolii плазмиды, кроме структурных генов нитрогеназы, содержат гены, отвечающие за развитие корневых клубеньков у определенных видов бобовых.
Конструирование плазмид, несущих nif-гены, позволяет передавать способность к фиксации азота организмам, не обладающим этим свойством. Среди бактерий, кроме Е. , такой перенос осуществлен для бактерий Salmonella typhimurium, Erwinia herbicola и других. Однако подобные манипуляции могут приводить к нежелательным эффектам. Так, перенос генов в штамм Erwinia (бактерии, вызывающие гниение растений) может усилить его патогенное действие. Кроме того, существует вероятность случайного переноса вместе с nif-генами каких-то нежелательных генов.
В настоящее время внимание ученых привлекают проблемы введения генов азотфиксации в клетки растений, создания ризоценозов между небобовыми растениями (особенно злаками) и азотфиксирующими организмами, повышения мощности корневой системы бобовых растений для увеличения на ней количества клубеньков. Кроме того, предполагается создание новых азотфиксирующих систем путем введения азотфиксирующих микроорганизмов в каллусные ткани растений с последущим образованием из них растений-регенерантов, а также повышение эффективности фиксации азота путем воздействия на гены, контролирующие этот процесс.
Наиболее интересна первая проблема — введение nif-генов в клетки растений. Однако ее решение сопряжено с рядом трудностей. Основная — разрушение нитрогеназы под воздействием кислорода. У азотфиксирующих микроорганизмов существует ряд приспособлений, защищающих бактерии от свободного кислорода. Среди них присутствие в клетках клубеньков лешглобина — гем- содержащего белка, который встраивается в мембрану бактероида (увеличенной в размере бактериальной клетки, характеризующейся наибольшей способностью к фиксации азота) и регулирует поступление кислорода. Легоглобин кодируется в геноме растительной клетки-хозяина, но его синтез начинается только после проникновения бактерий в эту клетку. У цианобактерий механизм защиты нитрогеназы от кислорода иной. Азотфиксация идет в гетероцистах, а фотосинтез — в обычных клетках. Поэтому кислород, выделяющийся в процессе фотосинтеза, не ингибирует фиксацию азота. Введение только nif -генов в какую-то растительную клетку не решает проблемы. Если нитрогеназа будет синтезироваться в этой клетке, в частности в клетках злаков, то она разрушится под действием кислорода, присутствующего в клетке. Кроме того, сама клетка, в которую переносят гены азотфиксации, может быть не приспособлена к синтезу и расходованию большого количества энергии, которое требуется для фиксации азота.
Таким образом, более перспективно повышение эффективности фиксации азота в уже существующих природных системах за счет воздействия на гены, контролирующие этот процесс, а также увеличение мощности корневой системы бобовых растений и создание новых азотфиксирующих систем с помощью методов клеточной инженерии.
5.6. Изменение качества плодов
В настоящее время найдены способы регулирования сроков созревания плодов с помощью методов генетической инженерии. Для этого используют метод создания антисмысловой РНК, который позволяет управлять работой интересующего гена. В растениях ген PG контролирует синтез полигалактуроназы — фермента, который участвует в разрушении пектина. Данный фермент синтезируется в период созревания плодов томата, они становятся мягкими, что значительно сокращает срок их хранения. Отключение гена PG позволяет получить растения с новыми свойствами — увеличенным сроком хранения и более высокой устойчивостью к грибным заболеваниям. Эту же технологию можно применить для регулирования активности другого гена — гена EFE, продуктом которого является фермент, участвующий в биосинтезе этилена — фитогормона, одной из функций которого служит ускорение созревания плодов. Отключение данного гена также может увеличить срок хранения плодов.
5.7. Устойчивость растений к гербицидам
В сельском хозяйстве широко используют гербициды — химические соединения, применяемые для уничтожения сорной растительности. Гербициды широкого спектра действия могут не только уничтожать сорняки, но и угнетать рост культурных растений. В связи с этим возникает необходимость в создании растений, устойчивых к данным веществам. Существует два подхода к решению этой проблемы: прямая селекция устойчивых к гербицидам мутантных форм растений, или мутантных клеточных штаммов (клеточная селекция), и генно-инженерный метод, который состоит во введении в растения генов гербицид-резистентности растительного или бактериального происхождения.
Изучение механизмов устойчивости служит основой для создания трансгенных растений. Оно включает четыре основных этапа: выявление мишеней действия гербицидов в клетке растений; отбор растений, устойчивых к данному гербициду в качестве источника генов резистентности; идентификация и клонирование этих генов; изучение их экспрессии для использования в трансгенных конструкциях.
Благодаря использованию методов генетической инженерии были созданы новые, устойчивые к различным гербицидам сельскохозяйственные культуры. В геном этих культур вводились мутантные гены, кодирующие синтез ферментов, на которые гербициды (атразин, бромоксилин, имидазол) не оказывают негативного действия. Например, растения лядвенца рогатого (Lotus corniculatus) были трансформированы с помощью штамма А281/рСВЕ21. Эта бактерия содержит плазмиду с встроенным геном bar, кодирующим фермент, придающий устойчивость к гербициду биалофосу. Трансгенные растения содержали ген bar и были невосприимчивы к гербициду (А. М. Стефанович, Г. Н. Ралдугина, 1999). Однако в тканях таких растений наблюдается накопление гербицидов, и использовать эти растения можно только в технических целях. Вместе с тем было показано, что введение генов, кодирующих другие ферменты, позволяет проводить детоксикацию гербицидов, создавая таким образом растения, пригодные в пищу. Так, детоксикация действующего вещества гербицида 2,4-D осуществляется при переносе в растение гена монооксигеназы, глифосата — при введении гена фосфонатазы, бромоксилина — гена нитрилазы.
5.8. Устойчивость растений к фитопатогенам и насекомым
Необходимость создания растений, устойчивых к фитопатогенам, связана с тем, что насекомые-вредители, вирусные и другие заболевания служат причиной значительного снижения урожайности сельскохозяйственных культур.
Наибольший урон растениям наносят грибные, бактериальные и вирусные патогены, насекомые-вредители. В растении существуют защитные механизмы, которые в большей или меньшей степени (в зависимости от устойчивости растений) начинают действовать в ответ на проникновение фитопатогенов в клетку. Во-первых, начинается синтез соединений, вызывающих гибель патогенов. Примером могут служить специфические белки PRP (pathogen related proteins). Из них наиболее изучены ферменты хитиназы и бета-1,3-глюконазы, которые угнетают рост грибов и некоторых видов бактерий, разрушая их клеточные стенки. Во-вторых, могут создаваться структурные барьеры, препятствующие распространению инфекции. Это достигается благодаря лигнификации клеточных стенок. Той же цели — защите клеток — служит присутствие в клеточных стенках белков-экстенсинов и олигосахаридов.
Применение методов генетической инженерии, использующих естественные защитные механизмы, позволяет получать трансгенные растения, устойчивые к грибной, бактериальной и вирусной инфекции. Так, гены хитиназы и глюконазы кодируются одиночными генами. Благодаря этому были получены трансгенные растения табака и турнепса, в состав генома которых ввели ген хитиназы. Лабораторные и полевые испытания выявили большую устойчивость трансгенных растений. В растения томатов был введен ген защитных пептидов редьки (дефензинов) rs, отвечающих за устойчивость к фитопатогенным грибам. Наконец, перспективны клонирование и перенос генов, кодирующих специфические белки (small antibiotic-like proteins), содержащиеся в семенах многих растений. Эти белки защищают семена в период покоя и во время прорастания от грибных и бактериальных инфекций.
Другой подход к получению трансгенных растений, устойчивых к вирусной инфекции, состоит во введении в геном исходных растений гена оболочки вируса. Это приводит к ингибированию размножения вируса и снижению инфицированности. Благодаря такому подходу был получен стойкий антивирусный эффект у растений табака, трансформированных геном оболочки вируса табачной мозаики (ВТМ).
Еще одна группа методов получения трансгенных растений, устойчивых к действию фитовирусов, включает введение и экспрессию генов антивирусных антител, вирусных сателлитных РНК. Интересный эффект дало введение в геном растений гена человеческого интерферона JFN — одного из ключевых белков индукции иммунитета у млекопитающих. С помощью вируса мозаики цветной капусты геном интерферона были трансформированы растения турнепса, табака, картофеля, что повысило устойчивость этих растений к вирусным заболеваниям.
Вполне традиционным для придания устойчивости к насеко- мым-вредителям стало внесение в растения гена Bt бактерий Bacillus thurengiensis. Этот ген отвечает за синтез белка протоксина, который при попадании в кишечник личинок насекомых разрушается. Продукт данного ферментативного расщепления — 5-эндотоксин — вызывает гибель насекомых. Интерес к этой группе белков вызван их широким спектром действия. Найдены новые классы эндотоксинов, которые токсичны к клещам, нематодам, одноклеточным паразитирующим микроорганизмам и т.д. Более того, в настоящее время сконструирован искусственный ген Bt, более эффективный, с более широким спектром устойчивости по сравнению с природным геном.
Другой путь повышения устойчивости растений к вредителям, в частности к фитопатогенным грибам, — это экспрессия в клетках растений генов — ингибиторов протеиназ. Для данной цели перспективно использование гетерологичных генов растений, так как многие ткани и органы растений содержат ингибиторы, специфически настроенные против протеиназ насекомых и грибов, но безвредных для человека и животных.
Еще одно направление создания трансгенных устойчивых растений связано с повышением их иммунитета. У устойчивых растений в ответ на атаку патогенов происходит изменение метаболизма: накапливаются перекись водорода, салициловая кислота, фитоалексины — соединения, выполняющие защитную роль в растении. Поэтому существенное увеличение с помощью изменения генома уровня салициловой кислоты и других соединений в ответ на проникновение фитопатогена может стать перспективным для получения устойчивых растительных организмов.
5.9. Устойчивость растений к абиотическим стрессам
Адаптация растений в природе и, следовательно, их способность к выживанию при неблагоприятных условиях среды обеспечиваются тремя способами. Во-первых, физиологические механизмы, позволяющие растениям избежать неблагоприятные воздействия (например, период покоя). Во-вторых, адаптация осуществляется благодаря морфологическим приспособлениям: толстому слою кутикулы на листьях, уменьшению листовой поверхности, ее опушению, которые предотвращают излишнюю потерю влаги растениями. В-третьих, негативное влияние внешней среды может быть преодолено с помощью изменений метаболизма. Именно этот последний адаптационный механизм наиболее доступен для генно-инженерных исследований. Например, известно, что при водном стрессе у высших растений основным защитным механизмом, связанным с изменением метаболизма, является накопление в клетках пролина, глицинбеатина и других осмопротекгоров.
Экспериментально было показано, что стрессовый ответ у бактерий и высших растений выражается сходно. И у растений, и у бактерий начинается усиленный синтез молекул осмопротекторов, механизм действия которых состоит в установлении осмотического баланса между цитоплазмой и окружающей средой, а также стабилизации белковых молекул. В бактериях биосинтез пролина хорошо изучен, известны гены, кодирующие ферменты этого процесса. Избирательная экспрессия генов осмопротекторов может привести к увеличению адаптационных качеств растения и, следовательно, к увеличению его продуктивности. Поэтому следующим шагом на пути создания устойчивых к стрессам растений было клонирование бактериальных генов, получение векторных конструкций на основе Ti-плазмиды и введение их в растения. Полученные трансгены синтезировали и накапливали пролин в 4—6 раз интенсивнее, чем обычные растения. Трансгенные побеги могли укореняться и расти при концентрации соли в среде 20 г/л (350 мМ).
У растений адаптация к низким температурам сопряжена с многочисленными физиологическими изменениями. При этом накапливаются растворимые вещества, понижающие осмотический потенциал клеток и уменьшающие вероятность образования крупных кристаллов льда. Кроме того, синтезируется большое количество белков с повышенным содержанием сульфгидрильных групп (-SH), которые обладают особо высокой способностью к гидратации, а гидратационная вода, как известно, практически не замерзает. Однако повышение устойчивости растений к замерзанию с помощью методов генной инженерии началось с изменения генома не растений, а бактерий. Исследователи Колорадского университета (США) выяснили, что повреждению растений при замерзании способствуют бактерии эпифитной (поверхностной) микрофлоры Pseudomonas syringae и Erwinia herbicola, белки которых служат центрами кристаллизации. Если обезвредить бактерии стрептомицином, то растения не замерзают при температуре -8 С. Но стрептомицин дорог и вреден, поэтому выгоднее было изменить генетику данного штамма бактерий, вырезав из генома определенный ген. Растения, инфицированные мутантным штаммом P. syringae, росли при отрицательной температуре. Однако оказалось, что бактерии мутантного штамма более живучи и способны вытеснить природный штамм, который, попадая в верхние слои атмосферы, способствует кристаллизации атмосферной влаги. Вероятно, уничтожение природного штамма могло бы привести к экологической катастрофе.
Следует отметить, что работы по генетической инженерии, возможности манипулирования генами растений представляют огромный интерес для фундаментальных исследований. Эти работы позволяют изучать основы молекулярной и клеточной биологии растительной клетки, глубинные механизмы процессов, происходящих в ней. Вместе с тем нельзя не задуматься о своевременности прикладного применения результатов генно-инженерных исследований.
Текст работы размещён без изображений и формул.
Полная версия работы доступна во вкладке «Файлы работы» в формате PDF
Первые трансгенные растения (растения табака со встроенными генами из микроорганизмов) были получены в 1983 г. Первые успешные полевые испытания трансгенных растений (устойчивые к вирусной инфекции растения табака) были проведены в США уже в 1986 г.
После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т.д. первые трансгенные продукты появились в продаже в США в 1994 г. Это были томаты Flavr Savr с замедленным созреванием, созданные фирмой «Calgen», а также гербицид-устойчивая соя компании «Monsanto». Уже через 1-2 года биотехнологические фирмы поставили на рынок целый ряд генетически измененных растений: томатов, кукурузы, картофеля, табака, сои, рапса, кабачков, редиса, хлопчатника.
В настоящее время получением и испытанием генетически модифицированных растений занимаются сотни коммерческих фирм во всем мире с совокупным капиталом более ста миллиардов долларов. В 1999 г. трансгенные растения были высажены на общей площади порядка 40 млн. га, что превышает размеры такой страны, как Великобритания. В США генетически модифицированные растения (GM Crops) составляют сейчас около 50% посевов кукурузы и сои и более 30-40% посевов хлопчатника. Это говорит о том, что генно-инженерная биотехнология растений уже стала важной отраслью производства продовольствия и других полезных продуктов, привлекающей значительные людские ресурсы и финансовые потоки. В ближайшие годы ожидается дальнейшее быстрое увеличение площадей, занятых трансгенными формами культурных растений.
Первая волна трансгенных растений, допущенных для практического применения, содержала дополнительные гены устойчивости (к болезням, гербицидам, вредителям, порче при хранении, стрессам).
Нынешний этап развития генетической инженерии растений получил название «метаболическая инженерия». При этом ставится задача не столько улучшить те или иные имеющиеся качества растения, как при традиционной селекции, сколько научить растение производить совершенно новые соединения, используемые в медицине, химическом производстве и других областях. Этими соединениями могут быть, например, особые жирные кислоты, полезные белки с высоким содержанием незаменимых аминокислот, модифицированные полисахариды, съедобные вакцины, антитела, интерфероны и другие «лекарственные» белки, новые полимеры, не засоряющие окружающую среду и многое, многое другое. Использование трансгенных растений позволяет наладить масштабное и дешевое производство таких веществ и тем самым сделать их более доступными для широкого потребления.
Создание гербицидоустойчивых растений
В новых, интенсивных сельскохозяйственных технологиях гербициды применяются очень широко. Это связано с тем. что на смену прежним экологически опасным гербицидам широкого спектра действия, обладающим токсичностью для млекопитающих и длительно сохраняющимся во внешней среде, приходят новые, более совершенные и безопасные соединения. Однако они обладают недостатком — подавляют рост не только сорняков, но и культурных растений Такие высокоэффективные гербициды, как, глифосат, атразины интенсивно изучаются на предмет выявления механизма толерантности к ним некоторых сорняков. Так, на полях, где широко используют атразин, довольно часто появляются атразинустойчивые биотипы у многих видов растении.
Изучение механизма устойчивости к гербицидам с целью получения методами генетической инженерии культурных растений, обладающих этим признаком, включает следующие этапы: выявление биохимических мишеней действия гербицидов в растительной клетке: отбор устойчивых к данному гербициду организмов в качестве источников генов устойчивости: клонирование этих генов: введение их в культурные растения и изучение их функционирования
Существуют четыре принципиально различных механизма, которые могут обеспечивать устойчивость к тем или иным химическим соединениям, включая гербициды: транспортный, элиминирующий, регуляционный и контактный. Транспортный механизм устойчивости заключается в невозможности проникновения гербицида в клетку. При действии элиминирующего механизма устойчивости вещества, попавшие внутрь клетки, могут разрушаться с помощью индуцируемых клеточных факторов, чаще всего деградирующих ферментов, а также подвергаться тому или иному виду модификации, образуя неактивные безвредные для клетки продукты. При регуляционной резистентности белок или фермент клетки, инактивирующийся под действием гербицида, начинает усиленно синтезироваться, ликвидируя таким образом дефицит нужного метаболита в клетке. Контактный механизм устойчивости обеспечивается изменением структуры мишени (белок или фермент), взаимодействием с которым связано повреждающее действие гербицида
Установлено, что признак гербицидоустойчивости является моногенным, то есть признак детерминируется чаще всего одним-единственным геном. Это очень облегчает возможность использования технологии рекомбинантной ДНК для передачи этого признака. Гены, кодирующие те или иные ферменты деструкции и модификации гербицидов, могут быть с успехом использованы для создания гербицидоустойчивых растении методами генетической инженерии.
Традиционные методы селекции создания сортов, устойчивых к гербицидам, очень, длительны и малорезультативны. Наиболее широко применяемый за рубежом гербицид глифосат (коммерческое название Roundup) подавляет синтез важнейших ароматических аминокислот, воздействуя на фермент 5-енолпирувилшикимат-З-фосфатсинтазу (ЕПШФ-синтаза). Известные случаи устойчивости к этому гербициду связаны либо с повышением уровня синтеза этого фермента (регуляционный механизм), либо с возникновением мутантного фермента, нечувствительного к глифосфату (контактный механизм). Из устойчивых к глифосфату растений был выделен ген ЕПШФ-синтазы и поставлен под промотор вируса мозаики цветной капусты. С помощью Ti-плазмиды эта генетическая конструкция была введена в клетки петунии. При наличии одной копии гена в регенерированных из трансформированных клеток растениях синтезировалось фермента в 20 — 40 раз больше, чем в исходных растениях, но устойчивость к глифосфату увеличилась только в 10 раз.
К числу наиболее распространенных гербицидов, используемых при обработке зерновых культур, относится атразин. Он подавляет фотосинтез, связываясь с одним из белков фотосистемы II и прекращая транспорт электронов. Устойчивость к гербициду возникает в результате точечных мутаций в этом пластохинон связывающем белке (замена серина на глицин), вследствие чего он теряет способность взаимодействовать с гербицидом. В ряде случаев удалось осуществить перенос гена мутантного белка в чувствительные к атразину растения с помощью Ti-плазмиды. Интегрированный в хромосому растений ген устойчивости был снабжен сигнальной последовательностью, которая обеспечивала транспорт синтезируемого белка в хлоропласты. Химерные растения проявляли значительную устойчивость к таким концентрациям атразина, которые вызывали гибель контрольных растений с геном белка дикого типа. Некоторые растения способны инактивировать атразин путем отщепления остатка хлора ферментом глутатион-S-трансфераза. Этот же фермент инактивирует и другие родственные гербициды триазинового ряда (пропазин, симазин и др.).
Существуют растения, естественная устойчивость которых к гербицидам основана на детоксикации. Так, устойчивость растений к хлорсульфурону может быть связана с дезактивацией молекулы гербицида путем его гидроксилирования и последующего гликозилирования введенной гидроксильной группы. Создание растений, устойчивых к патогенам и вредителям Устойчивость растений к тем или иным патогенам чаще всего является сложным мультигенным признаком.
Одновременная передача нескольких локусов трудна даже методами генной инженерии, не говоря о классических методах селекции. Более простым является другой путь. Известно, что у устойчивых растений при атаке патогенов изменяется метаболизм. Накапливаются такие соединения, как Н2О2, салициловая кислота, фитоаллексины. Повышенный уровень этих соединений способствует противостоянию растения в борьбе с патогенами.
Вот один из примеров, доказывающий роль салициловой кислоты в иммунном ответе растений. Трансгенные растения табака, которые содержат бактериальный ген, контролирующий синтез салицилат гидролазы (этот фермент разрушает салициловую кислоту), были неспособны к иммунному ответу. Поэтому изменение генно-инженерным путем уровня салициловой кислоты или выработки в растениях в ответ на патоген Н2О2 может быть перспективным для создания устойчивых трансгенных растений.
В фитовирусологии широко известен феномен индуцированной перекрестной устойчивости растений к вирусным инфекциям. Сущность этого явления состоит в том, что заражение растения одним штаммом вируса предотвращает последующую инфекцию этих растений другим вирусным штаммом. Молекулярный механизм подавления вирусной инфекции пока неясен. Показано, что для иммунизации растений достаточно введения отдельных вирусных генов, например генов капсидных белков. Так, ген белка оболочки вируса табачной мозаики перенесли в клетки табака и получили трансгенные растения, у которых 0,1% всех белков листьев был представлен вирусным белком. Значительная часть этих растений при инфицировании вирусом не проявляла никаких симптомов заболевания. Возможно, что синтезирующийся в клетках белок оболочки вируса мешает вирусной РНК нормально функционировать и формировать полноценные вирусные частицы. Установлено, что экспрессия капсидного белка вируса табачной мозаики, вируса мозаики люцерны, вируса огуречной мозаики, Х-вируса картофеля в соответствующих трансгенных растениях (табак, томаты, картофель, огурцы, перцы) обеспечивает высокий уровень их защиты от последующей вирусной инфекции. Причем у трансформированных растений не отмечалось снижения фертильности, нежелательного изменения ростовых и физиологических характеристик исходных экземпляров и их потомства. Полагают, что индуцированная устойчивость растений к вирусам обусловлена особым антивирусным белком, очень похожим на интерферон животных. Представляется возможным методом генетической инженерии усилить экспрессию гена, кодирующего этот белок, путем его амплификации или подстановки под более сильный промотор.
Следует отметить, что использование генетической инженерии для защиты растений от различных патогенных микроорганизмов в значительной мере сдерживается недостаточностью знаний о механизмах защитных реакций растений. Для борьбы с насекомыми-вредителями в растениеводстве используются химические средства — инсектициды. Однако они оказывают вредное влияние на млекопитающих, убивают и полезных насекомых, загрязняют окружающую среду, дороги, и кроме того, насекомые довольно скоро приспосабливаются к ним. Известно более 400 видов насекомых, устойчивых к используемым инсектицидам. Поэтому все большее внимание привлекают биологические средства борьбы, обеспечивающие строгую избирательность действия и отсутствие адаптации вредителей к применяемому биопестициду.
Уже довольно давно известна бактерия Bacillus thuringiensis, продуцирующая белок, являющийся очень токсичным для многих видов насекомых, в то же время безопасный для млекопитающих. Белок (дельта-эндотоксин, CRY-белок) продуцируется различными штаммами В. thuringiensis. Взаимодействие токсина с рецепторами строго специфично, что усложняет подбор комбинации токсин—насекомое. В природе найдено большое количество штаммов В. thuringiensis, чьи токсины действуют только на определенные виды насекомых. Препараты В. thuringiensis в течение десятилетий использовали для контроля насекомых на полях. Безопасность токсина и его составных белков для человека и других млекопитающих полностью доказана. Встраивание гена этого белка в геном растений дает возможность получить трансгенные растения, не поедаемые насекомыми.
Кроме видоспецифичности по действию на насекомых встраивание прокариотических генов дельта-токсинов в геном растений даже под контролем сильных эукариотических промоторов не привело к высокому уровню экспрессии. Предположительно такое явление возникло в связи с тем, что эти бактериальные гены содержат значительно больше адениновых и тиминовых нуклеотидных оснований, чем растительная ДНК. Эта проблема была решена путем создания модифицированных генов, где из природного гена вырезали и добавляли те или иные фрагменты с сохранением доменов, кодирующих активные части дельта-токсина. Так, например, с помощью таких подходов был получен картофель, устойчивый к колорадскому жуку. Получены трансгенные растения табака, способные синтезировать токсин. Такие растения были нечувствительны к гусеницам Manduca sexta. Последние погибали в течение 3 суток контакта с токсинпродуцирующими растениями. Токсинообразование и обусловленная им устойчивость к насекомым передавалась по наследству как доминантный признак.
В настоящее время так называемые Bt-растения (от В. thuringiensis) хлопка и кукурузы занимают основную долю в общем объеме генетически модифицированных растений этих культур, которые выращивают на полях США.
В связи с возможностями генной инженерии конструировать энтомопатогенные растения на основе токсина микробного происхождения еще больший интерес к себе вызывают токсины растительного происхождения. Фитотоксины являются ингибиторами белкового синтеза и осуществляют защитную функцию, направленную против насекомых-вредителей микроорганизмов и вирусов. Лучше всех среди них изучен рицин, синтезируемый в клещевине: его ген клонирован и установлена нуклеотидная последовательность. Однако высокая токсичность рицина для млекопитающих ограничивает генноинженерные работы с ним только техническими культурами, не используемыми в пищу человека и на корм животным. Токсин, вырабатываемый фитолаккой американской, эффективен против вирусов и безвреден для животных. Механизм его действия заключается в инактивации собственных рибосом при проникновении в клетки различного рода патогенов, в том числе фитовирусов. Пораженные клетки некротизируются, предотвращая размножение патогена и его распространение по растению. В настоящее время проводятся исследования по изучению гена этого белка и передаче его в другие растения.
Вирусные болезни широко распространены среди насекомых, поэтому для борьбы с насекомыми-вредителями можно использовать природные вирусы насекомых, препараты которых называют вирусными пестицидами. В отличие от ядохимикатов они обладают узким спектром действия, не убивают полезных насекомых, они быстро разрушаются во внешней среде и не опасны для растений и животных. Наряду с вирусами насекомых используются как биопестициды некоторые грибы, поражающие насекомых-вредителей. Применяемые сейчас биопестициды являются природными штаммами энтомопатогенных вирусов и грибов, однако не исключена возможность создания в будущем методами генетической инженерии новых эффективных биопестицидов.
Повышение устойчивости растений к стрессовым условиям
Растения очень часто подвергаются воздействию различных неблагоприятных факторов окружающей среды: высокие и низкие температуры, недостаток влаги, засоление почв и загазованность среды, недостаток или, напротив, избыток некоторых минеральных веществ и т. д. Этих факторов множество, поэтому и способы защиты от них многообразны — от физиологических свойств до структурных приспособлений, позволяющих преодолевать их пагубное действие.
Устойчивость растений к тому или иному стрессовому фактору является результатом воздействия множества разных генов, поэтому говорить о полной передаче признаков толерантности от одного вида растения другому генноинженерными методами не приходится. Тем не менее у генетической инженерии имеются определенные возможности для повышения устойчивости растений. Это касается работы с отдельными генами, контролирующими метаболические ответы растений на стрессовые условия, например сверхпродукцию пролина в ответ на осмотический шок, на действие засоления, синтез особых белков в ответ на тепловой шок и т. д. Дальнейшее углубленное изучение физиологической, биохимической и генетической основы ответной реакции растения на условия среды, несомненно, позволит применять методы генетической инженерии для конструирования устойчивых растений.
Пока можно отметить лишь косвенный подход для получения морозоустойчивых растений, основанный на генноинженерных манипуляциях с Pseudomonas syringae. Этот микроорганизм, сосуществующий с растениями, способствует их повреждению ранними заморозками Механизм явления связан с тем, что клетки микроорганизма синтезируют особый белок, локализующийся во внешней мембране и являющийся центром кристаллизации льда. Известно, что формирование льда в воде зависит от веществ, могущих служить центрами образования льда. Белок, вызывающий формирование кристаллов льда в различных частях растения (листья, стебли, корни), является одним из главных факторов, ответственных за повреждение тканей растений, чувствительных к ранним заморозкам. Многочисленные эксперименты в строго контролируемых условиях показали, что стерильные растения не повреждались заморозками вплоть до —6—8° С, тогда как у растений, имеющих соответствующую микрофлору, повреждения возникали уже при температурах —1,5—2° С. Мутанты этих бактерий, потерявшие способность синтезировать белок, вызывающий формирование кристаллов льда, не повышали температуру образования льда, и растения с такой микрофлорой были устойчивы к заморозкам. Штамм таких бактерий, распыленный над клубнями картофеля, конкурировал с обычными бактериями, что приводило к повышению морозоустойчивости растений. Возможно, такие бактерии, созданные с помощью методов генной инженерии и используемые в качестве компонента внешней среды, будут служить для борьбы с заморозками.
Повышение эффективности биологической азотфиксации
Хорошо изучен фермент ответственный за восстановление молекулярного азота до аммония. — нитрогеназа. Структура нитрогеназы одинакова у всех азотфиксирующих организмов. При фиксации азота непременным физиологическим условием является защита нитрогеназы от разрушения под действием кислорода. Лучше всех среди азотфиксаторов изучены ризобии, образующие симбиоз с бобовыми растениями, и свободноживущая бактерия Klebsiella pneumoniae. Установлено, что у этих бактерий за фиксацию азота ответственно 17 генов — так называемых nif-генов. Все эти гены сцеплены друг с другом и расположены в хромосоме между генами ферментов биосинтеза гистидина и генами, определяющими усвоение шикимовой кислоты. У быстрорастущей ризобии nif-гены существуют в форме мегаплазмиды, содержащей 200—300 тысяч пар нуклеотидов.
Среди генов азотфиксации выявлены гены, контролирующие структуру нитрогеназы, белковый фактор, принимающий участие в транспорте электронов, регуляторные гены. Регуляция генов азотфиксации довольно сложна, поэтому генноинженерный перенос азотфиксирующей функции от бактерий непосредственно высшим растениям в настоящее время уже не обсуждается. Как показали эксперименты, даже в самом простом эукариотическом организме — дрожжах не удалось добиться экспрессии nif-генов, хотя они и сохранялись в течение 50 генераций.
Эти опыты показали, что диазотрофность (азот-фиксация) свойственна исключительно прокариотическим организмам, и nif-гены не смогли преодолеть барьер, разделяющий прокариоты и эукариоты, из-за слишком сложной своей структуры и регуляции генами, расположенными вне nif-области. Возможно, более удачным окажется перенос nif-генов с помощью Ti-плазмид в хлоропласты, поскольку механизмы экспрессии генов в хлоропластах и в клетках прокариот близки. В любом случае нитрогеназа должна быть защищена от ингибирующего действия кислорода. Кроме того, фиксация атмосферного азота — очень энергоемкий процесс. Вряд ли растение под влиянием nif-генов может так кардинально изменить свой метаболизм, чтобы создать все эти условия. Хотя не исключено, что в будущем методами генетической инженерии можно будет создать более экономно работающий нитрогеназный комплекс.
Более реально использование генноинженерных методов для решения следующих задач: повышение способности ризобии колонизировать бобовые растения, повышение эффективности фиксации и ассимиляции азота путем воздействия на генетический механизм, создание новых азотфиксирующих микроорганизмов путем введения в них nif-генов, передача способности к симбиозу от бобовых растений к другим.
Первостепенной задачей генетической инженерии для повышения эффективности биологической фиксации азота является создание штаммов ризобии с усиленной азотфиксацией и колонизирующей способностью. Колонизация бобовых растений ризобиями протекает очень медленно, лишь единичные из них дают начало клубенькам. Это происходит потому, что местом инвазии ризобии является только одна небольшая область между точкой роста корня и ближайшим к ней корневым волоском, находящимся на стадии формирования. Все остальные части корня и развившиеся корневые волоски растения нечувствительны к колонизации. В ряде случаев сформировавшиеся клубеньки оказываются неспособными фиксировать азот, что зависит от многих растительных генов (выявлено не менее пяти), в частности от неблагоприятного сочетания двух рецессивных генов.
Традиционными методами генетики и селекции удалось получить лабораторные штаммы ризобий с более высокой колонизирующей способностью. Но они в полевых условиях испытывают конкуренцию со стороны местных штаммов. Повышение их конкурентоспособности, видимо, можно осуществить генноинженерными методами. Повышение эффективности процесса азотфиксации возможно применением генноинженерных приемов, основанных на увеличении копий гена, усилении транскрипции тех генов, продукты которых образуют «узкое» место в каскадном механизме азотфиксации, путем введения более сильных промоторов и т. п. Важно повышение коэффициента полезного действия самой нитро-геназной системы, осуществляющей непосредственное восстановление молекулярного азота в аммиак.
Повышение эффективности фотосинтеза
С4-растения характеризуются высокими темпами роста и скоростью фотосинтеза, у них практически отсутствует видимое фотодыхание. У большинства сельскохозяйственных культур, относящихся к С3-растениям, высокая интенсивность фотодыхания. Фотосинтез и фотодыхание — тесно связанные процессы, в основе которых лежит бифункциональная активность одного и того же ключевого фермента — рибулозобисфосфат-карбоксилазы (РуБФК). РуБФ-карбоксилаза может присоединять не только С02, но и 02, то есть осуществляет реакции карбоксилирования и оксигенирования. При оксигенировании РуБФ образуется фосфогликолат, который служит основным субстратом фотодыхания — процесса выброса С02 на свету, в результате чего теряется часть фотосинтетических продуктов. Низкое фотодыхание у С4-растений объясняется не отсутствием ферментов гликолатного пути, а ограничением оксигеназной реакции, а также реассимиляцией С02 фотодыхания.
Одной из задач, стоящих перед генетической инженерией, является исследование возможности создания РуБФК с преобладающей карбоксилазной активностью.
Получение растений с новыми свойствами
В последние годы ученые используют новый подход для получения трансгенных растений с «antisense RNA» (перевернутой или антисмысловой РНК), который позволяет управлять работой интересуемого гена. В этом случае при конструировании вектора копию ДНК (к-ДНК) встраиваемого гена переворачивают на 180°. В результате в трансгенном растении образуется нормальная молекула мРНК и перевернутая, которая в силу комплементарности нормальной мРНК образует с ней комплекс и закодированный белок не синтезируется.
Такой подход использован для получения трансгенных растений томатов с улучшенным качеством плодов. Вектор включал к-ДНК гена PG, контролирующего синтез полигалактуроназы — фермента, участвующего в разрушении пектина, основного компонента межклеточного пространства растительных тканей. Продукт гена PG синтезируется в период созревания плодов томатов, а увеличение его количества приводит к тому, что томаты становятся более мягкими, что значительно сокращает срок их хранения. Отключение этого гена в трансгенах позволило получить растения томатов с новыми свойствами плодов, которые не только значительно дольше сохранялись, но и сами растения были более устойчивы к грибным заболеваниям.
Такой же подход можно применить для регулирования сроков созревания томатов, а в качестве мишени в этом случае используют ген EFE (ethylene-forming enzyme), продуктом которого является фермент, участвующий в биосинтезе этилена. Этилен — это газообразный гормон, одной из функций которого является контроль за процессом созревания плодов.
Cтратегия антисмысловых конструкций широко применима для модификации экспрессии генов. Эта стратегия используется не только для получения растений с новыми качествами, но и для фундаментальных исследований в генетике растений. Следует упомянуть еще об одном направлении в генной инженерии растений, которое до недавнего времени в основном использовали в фундаментальных исследованиях — для изучения роли гормонов в развитии растений. Суть экспериментов заключалась в получении трансгенных растений с комбинацией определенных бактериальных гормональных генов, например только iaaM или ipt т.д. Эти эксперименты внесли существенный вклад в доказательство роли ауксинов и цитокининов в дифференцировке растений.
В последние годы этот подход стали использовать в практической селекции. Оказалось, что плоды трансгенных растений с геном iaaM, находящимся под промотором гена Def (ген, который экспрессируется только в плодах), являются партенокарпическими, то есть сформировавшимися без опыления. Партенокарпические плоды характеризуются либо полным отсутствием семян, либо очень небольшим их количеством, что позволяет решить проблему «лишних косточек», например в арбузе, цитрусовых и т.д. Уже получены трансгенные растения кабачков, которые в целом не отличаются от контрольных, но практически не содержат семян.
Обезоруженную, лишенную онкогенов Ti-плазмиду ученые активно используют для получения мутаций. Этот метод носит название Т-ДНК-инсерционного мутагенеза. Т-ДНК, встраиваясь в геном растения, выключает ген, в который она встроилась, а по утрате функции можно легко отбирать мутанты (явление сайлесинга – замолкания генов). Этот метод замечателен также тем, что позволяет сразу обнаружить и клонировать соответствующий ген. В настоящее время таким способом получено множество новых мутаций растений и соответствующие гены клонированы. М. А. Раменской на основе Т-ДНК мутагенеза получены растения томатов с неспецифической устойчивостью к фитофторозу. Не менее интересен и другой аспект работ — получены трансгенные растения с измененными декоративными свойствами. Один из примеров — это получение растений петунии с разноцветными цветками. На очереди голубые розы с геном, контролирующим синтез голубого пигмента, клонированным из дельфиниума.
Зачем все это?
Стоит задавать этот вопрос перед началом любого более-менее осмысленного дела. А если дело — выведение генетически уникального организма, то и подавно. Хотите вывести розы, что сражают ядами — блокаторами дыхательной цепи каждого, кто укололся об их шипы, — да будет так. Подумываете вывести репу, без рыб рыбий жир производящую, — мы только поддержим. Поскольку редакция наша за мир во всем мире, остановимся на варианте с репкой.
Чьи гены пересаживать будем?
Чтобы репка наша рыбий жир производила, можно попробовать пересадить ей ген лосося, что кодирует белок, что омега-3 жирные кислоты* производит. Но, вполне возможно, у лососей производство полезного жира зависит от целого каскада белков и пересаживать придется сразу десяток генов. А возможно, и белки лосося несовершенно будут работать — слишком уж разная биохимия. Для поиска идеального донора генов придется прошерстить мириады страниц научных статей. И, как часто бывает, кто ищет, тот находит. Известны науке растения такие, что омега-3 жирные кислоты производят. Есть об этом статья, а в ней пишут, что у некоторых примул (широко известных под именем первоцвет) в цветках встречаются два белка, ответственных за производство омега-3 жирных кислот. Известны и гены, что кодируют в примулах эти белки. И встречаются эти гены еще у грибов рода нейроспора. Но примула репке более близкая родня, оттого и донором первостепенным ее выбрать стоит.
*Омега-3 ненасыщенные жирные кислоты — этим длинным словосочетанием химики называют органические кислоты, имеющие двойную связь после третьего с конца атома углерода. Омега-3 кислоты могут иметь двойные связи и в других позициях, тогда их называют полиненасыщенными. Эти жирные кислоты необходимы для здорового роста, развития и функционирования нашего организма. Их недостаток связывают с развитием клинической депрессии, отложением холестериновых бляшек, а регулярное потребление — с увеличением продолжительности жизни. Наши собственные клетки вырабатывать омега-3 жирные кислоты не умеют, поэтому их необходимо употреблять с пищей. В большом количестве они содержатся в жире морской рыбы.
Экспедиция за генами
Донор генов для репки, что веганов мира всего рыбьим жиром нерыбьего происхождения снабдит, обитает в предгорьях Кордильер. Придется отправиться за генами в путешествие. Только не спешите паковать чемоданы — сначала посадите репку, чтобы было куда гены потом пересаживать. Посадили? Отлично, можно лететь за примулой-донором. Не забудьте вооружиться канцелярским дыроколом, маленькими стерильными пробирками-эппендорфами и 70%-ным раствором спирта. Этот набор можно назвать «starter pack юного генного инженера». Глобальная база биоразнообразия GBIF* расскажет нам, где донор генов прячется, где и когда его в последний раз заметили, — начните поиски с этой локации. И, о чудо — удача с первой попытки! Перед вами заветный первоцвет. Не теряйте самообладания: твердой рукой берите дырокол и хладнокровно дырявьте им лист. В листе осталась дырка, а в дыроколе — кружочек. Этот кружочек нужно положить в ту маленькую пробирку, что с собой захватили, да залить все спиртом.
*GBIF — база данных, в которой зафиксировано, какой вид организмов где и когда был встречен. С помощью GBIF можно найти хоть кашалота, хоть иерихонскую розу.
Три волшебные буквы
Отобранный кружочек содержит несколько тысяч клеток. В каждой клетке — по копии заветного гена. Только как его оттуда извлечь? Для этого генетический материал отправляют на три нашумевших буквы — ПЦР*. Реакция эта позволяет сделать мириады копий любого фрагмента ДНК, нужно лишь правильно ингредиенты подобрать. Кидаем в котел правильно подобранные праймеры, фермент-полимеразу да нуклеотидов запас. Последний штрих — тщательно измельчаем кружочек из дырокола и отправляем в ПЦР-котел. Там происходит магия: праймер ищет в геноме донора комплементарный участок и связывается с ним, образуя этакую взлетно-посадочную полосу для ДНК-полимеразы. Полимераза видит праймер, садится на него и начинает клепать копию вожделенного гена. На разных стадиях процесса вы то нагреваете, то охлаждаете пробирку, в которой творится эта молекулярная алхимия.
*ПЦР — полимеразно-цепная реакция. Изобретение 1980-х, которое принесло своему автору Нобелевскую премию по химии. Эта реакция — один из самых быстрых и эффективных способов получить неограниченно большое количество копий нужного гена. Схема реакции достаточно проста. Сначала необходимо подобрать участок ДНК, начиная с которого необходимо «копировать» фрагмент ДНК и синтезировать праймер — короткий одноцепочечный фрагмент ДНК, комплементарный (помните школьную мантру А-Т, Г-Ц?) этому участку-мишени. С помощью праймера фермент ДНК-полимераза, отвечающий за копирование ДНК, «понимает», какой именно ген в длинной цепочке служит ее целью. Для наглядности можно представить, что ДНК — большая книга, ген — небольшой параграф, праймер — закладка, а ДНК-полимераза — копировальный аппарат. Не хватает только чернил (если угодно, бумаги), которые и составят копию. Ими служат нуклеотиды — структурные единицы (те самые А, Т, Г и Ц), буквы той самой ДНК-книги, копию фрагмента которой нужно сделать с помощью ПЦР.
Волшебная палочка
Теперь настало время достать волшебную палочку. Не бузинную, а палочковидную бактерию Agrobacterium tumefaciens — главного союзника генного инженера. Строго говоря, эта бактерия тоже генный инженер. Она умеет встраивать в геном растений гены из своей плазмиды*, что вызывает образование опухолей, клетки которых производят питательные для этих бактерий вещества. Ученые научились подсовывать агробактериям в плазмиду гены, которые сами хотят встроить в растения. С помощью набора ферментов перекраиваем плазмиду: вставляем туда наработанный из примулы ген, подшиваем пару генов, обеспечивающих устойчивость к антибиотикам у бактерий, и еще один — устойчивость к третьему антибиотику у растений. После молекулярной кройки делаем множество копий нашей плазмиды с помощью кишечной палочки E. coli** (они это делают почти сами и весьма охотно). Затем подсовываем плазмиды тем клеткам Agrobacterium tumefaciens, что не успели обзавестись собственными плазмидами. Убиваем антибиотиками, резистентность к которым зашивали в плазмиду, бактерий, коих не удалось модифицировать. В результате выживут только устойчивые — те, которым пересадили ген рыбьего жира. Дальше можно пересаживать его в репку.
*Плазмида — это небольшая молекула ДНК, обособленная от основного генетического материала, заключенного в хромосоме. У людей их нет, а вот у бактерий частенько встречаются.
**E. coli — излюбленный питомец микробиологов, биотехнологов, генетиков и представителей других отраслей биологии. Именуется эшерихией (лат. Escherichia) в честь Теодора Эшериха, микробиолога из тогда еще Австро-Венгрии. Сам Эшерих выделил ее из фекалий здоровых людей и окрестил Bacterium coli, то есть кишечной палочкой в переводе с латыни. За пределами научного мира ее до сих пор так частенько называют.
Пересадка генов
У нас был донор гена — ХХХ. Вектором-посредником мы выбрали агробактерий. Теперь настала пора подготовить реципиента — репку. За время путешествий и молекулярных манипуляций она успела вырасти. Но нельзя просто так взять и пересадить ген в репку. Нужно выделить из нее каллус. Так называются стволовые клетки растений. Для этого берем стерильный скальпель и надрезаем стебель. Вскоре на нем появится зеленоватая вязкая жидкость — это и есть каллус. Его надо пересадить на питательную среду. А затем подсадить к культуре каллуса еще и агробактерий. И сделать это 20 раз подряд: высадить 20 чашек каллуса, да чтобы погуще, и подсадить бактерий во все. В засеянных чашках Петри рождается страсть, которой лучше не мешать.
Выждали. Если работали не слишком стерильно, то в некоторых чашках заведется плесень или еще чего странное — на утилизацию. Оставшиеся, с виду не заросшие чашки Петри обрабатываем всеми тремя антибиотиками. В нескольких чашках Петри все опять погибает — на утилизацию. Но в паре чашек жизнь продолжает теплиться — там-то, видимо, генная модификация прошла успешно. Пересаживаем трансгенные стволовые клетки на новые и новые чашки Петри, заставляем их делиться, а после трансформироваться в полноценное растение. Для этого им в чашку нужно подмешать немного гормонов — ауксинов и цитокининов*.
Вот потихоньку из бесформенной клеточной массы начинает вытягиваться трансгенной репы росток. Будет он большой да сильный, и рыбьего жира в нем станет и на дедку, чтобы сердечно-сосудистыми не болел, и на бабку, чтобы нервная система была в порядке, и болявой внучке для иммунитета. А жучка ГМ-репу вряд ли оценит, ибо хищница.
*Ауксины и цитокинины — у растений, как и у животных, существует гормональная регуляция. Две основные группы фитогормонов — это ауксины и цитокинины. Их действие сложно и многогранно, особенно в различных комбинациях, но если сильно упрощать, то первые (то бишь ауксины) способствуют апикальному росту (то есть побегов вертикально вверх, а корней — вниз), а вторые (то бишь цитокинины) росту вширь. Чтобы вырастить из аморфной массы трансгенных стволовых клеток нечто похожее на репку, клеткам нужно подсказывать, как и в каком направлении им делиться. Гормоны и будут посредником в этих переговорах.
Сказки сказками, но в магазинах западных уже продают наборы для домашней генной инженерии. В них, как правило, включают и компактные центрифуги, вращающиеся с частотой 10 тысяч оборотов в минуту, и приборы для ПЦР с контролем температуры, и автоматические пипетки с набором разовых наконечников, и чашки для выращивания клеток, и культуры бактерий-векторов… И это лишь первые несколько позиций из огромного перечня оборудования, что включают эти наборы. Ценник, кстати, не слишком сказочный. Собрать домашнюю лабораторию обойдется в тысячу-другую долларов.